
基質と基質結合分子の解析を通して、紡錘体形成メカニズムを明らかにする
M期キナーゼ(mitotic kinase)と総称されるセリン/スレオニンキナーゼはM期進行を制御する重要なキナーゼ群です。私たちは中でもPlk1に注目しています。Plk1は酵母からヒトまで高度に保存されており、M期における重要性や癌化との関連が明らかになっているものの、基質分子がまだ少数しか同定されていません。私たちは固相リン酸化法を用いた基質候補分子のスクリーニングを行い、50以上の候補を得ています。

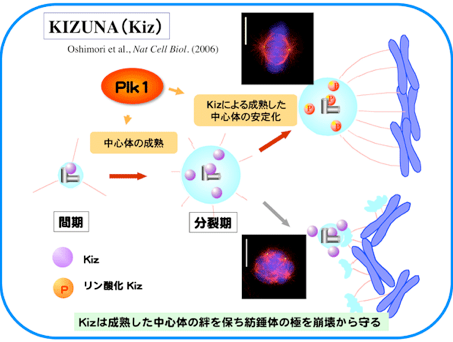
更に私たちは最近、Kizunaに結合しKizunaの中心体局在を担う因子としてCep72を同定しました。Cep72はKizunaだけではなく、CG-NAP/AKAP450 やgamma-tubulin ring complexなど他の主要な中心体構成要素の一部を中心体に局在させる働きがありました。特にCG-NAPをPCMに局在させることは中心体の微小管重合中心(MTOC)活性に必須であり、また分裂期には染色体を中期板に整列させることが可能な紡錘体を形成するために必須であることを示しました (Oshimori N., et al. EMBO J., 28:2066-2076, 2009)。
KizunaとCep72の更なる解析から、新たな中心体、紡錘体極の機能・構造制御機構の解明が期待できます。
