中心体で微小管重合を制御する分子
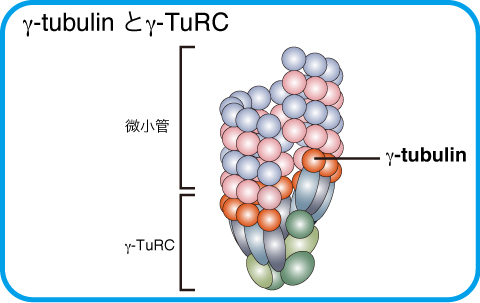
γ-tubulinは酵母からヒトまで真核生物で広く保存された中心体に局在するタンパク質です。哺乳類ではGCP2, GCP3などのGCPタンパク質とともにγ-tubulin環状複合体 (γ-tubulin ring complex, γ-TuRC)を形成し、微小管の重合核として働いています。
哺乳動物に存在する2種類のγ-tubulin
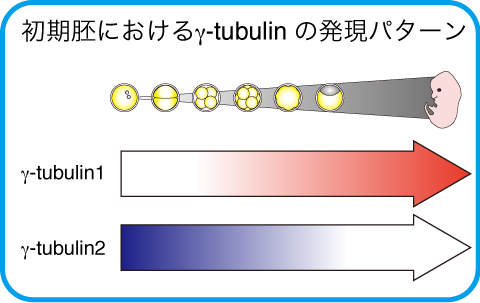
哺乳類を除く脊椎動物ではγ-tubulinの遺伝子は1つしかありませんが、哺乳類には2種類のγ-tubulin遺伝子が存在します。γ-tubulin1とγ-tubulin2は非常に相同性が高く、どちらもγ-TuRC複合体を形成し、中心体に局在することから、その機能の違いについては、全く解明されていません。
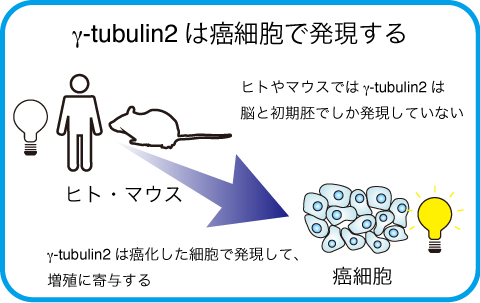
マウスの研究から、γ-tubulin1はユビキタスに発現していますが、γ-tubulin2は初期胚と脳に特異的に発現し、その他の組織では発現していないことが報告されています。ヒトでも同様の発現パターンだと考えられますが、これまでヒトの細胞や組織で、γ-tubulin1とγ-tubulin2をタンパク質レベルで区別して検出することが困難で、特にγ-tubulin2の発現を検出することが困難でした。
γ-tubulin2は癌細胞株で異所的に発現する
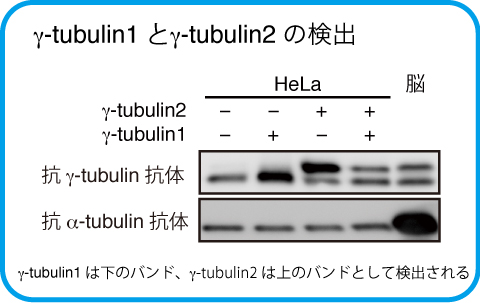
そこで私たちは、新たにヒトγ-tubulin1とγ-tubulin2をウエスタンブロッティングにより区別して検出できる方法を開発しました。SDS-PAGEで使用するアクリルアミド溶液に含まれるビスアクリスアミドの割合を下げた溶液で作製したゲル(Low-bis Gel)で電気泳動を行うと、γ-tubulin1とγ-tubulin2は異なる泳動度を示すバンドとして検出されます。その結果、共通するアミノ酸配列に対する抗体を用いることでγ-tubulin1とγ-tubulin2の発現量を定量すること可能になりました。
さらに、様々な癌細胞株についてγ-tubulin1と2の発現を解析すると、一部の癌細胞株では、生殖細胞や神経細胞由来ではないにも関わらず、γ-tubulin2の発現が認められ、またその発現が細胞の増殖に寄与していることを見出しました。 (Ohashi, T., et al., FEBS Lett., 590:1053-1063, 2016)
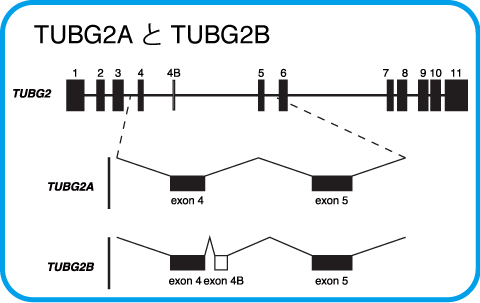
γ-tubulin2のmRNAには新規スプライシングバリアントが存在する
γ-tubulin2の遺伝子(TUBG2)は11のエキソンより構成されていますが、私たちは癌細胞株から、このmRNAに27ntの挿入配列を含む新規スプライシングバリアント(TUBG2Bと命名)を同定しました。
TUBG2Bの挿入配列は、TUBG2のエキソン4とエキソン5の間に位置するイントロン4領域に位置し、この領域が新規のエキソンとしてスプライシングを受けると考えられます。挿入配列が存在するイントロン4は、霊長類で特異的に長く、その他の哺乳動物と比べて約4倍の長さがあり、そのためTUBG2Bは霊長類で特異的に発現していると考えられます。
TUBG2Bの転写産物は複数の癌細胞株だけでなく、正常組織である脳からも検出されました。しかし、タンパク質としての発現は検出できず、また発現していたとしてもγ-tubulin環状複合体としての機能はないことを明らかにしました。(Ohashi, T., et al., FEBS Lett., 590:1053-1063, 2016)