微小管のプラス端の重合/脱重合を制御するモーター分子
MKif18Aはkinesin-8ファミリーに属するモーター分子です。微小管上をプラス端に向かって動くモーター活性と共に、微小管のプラス端特異的に重合を抑制する活性をもつという特徴があります。Kif18A は分裂期紡錘体の長さ、染色体整列、染色体オシレーション運動の制御に重要な分子ですが、Kif18A自身がどのような制御を受けているかについてはまだほとんど解明されていません。
MKif18Aはkinesin-8ファミリーに属するモーター分子です。微小管上をプラス端に向かって動くモーター活性と共に、微小管のプラス端特異的に重合を抑制する活性をもつという特徴があります。Kif18A は分裂期紡錘体の長さ、染色体整列、染色体オシレーション運動の制御に重要な分子ですが、Kif18A自身がどのような制御を受けているかについてはまだほとんど解明されていません。
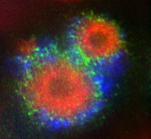
Kif18Aに関する興味深い現象のひとつに、その特徴的な細胞内局在変化があげられます。間期には核内に局在していますが、核膜崩壊とともに紡錘体微小管上に局在するようになります。前中期の初期には紡錘体微小管全体にほぼ一様に存在しますが、中期になると動原体に結合している動原体微小管(キネトコアファイバー)のプラス端に強く集積します。なぜ動原体微小管にだけ集積するのか、なぜ中期になると強く集積するのか。私たちはこの疑問点にアプローチし、微小管のプラス端の安定化がKif18Aのプラス端集積を引き起こすことを見出しました(Masuda N., and Shimodaira T., et al., Cell Struct. Funct., 2, 261-267, 2011)。
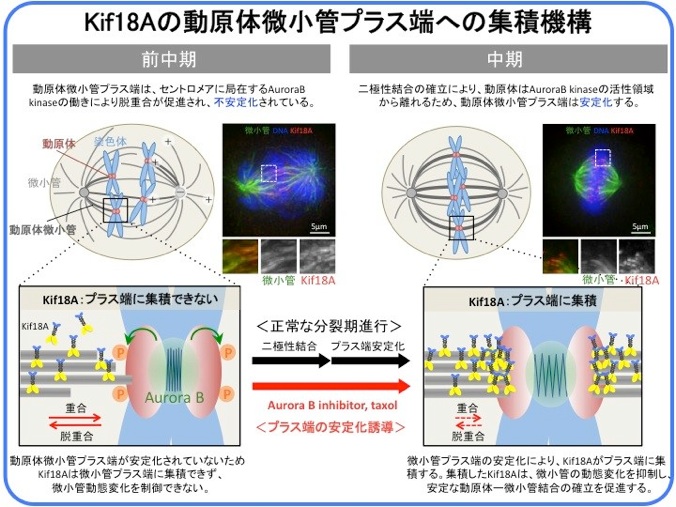
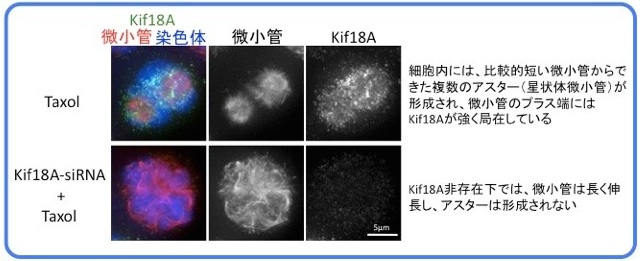
染色体が動原体微小管によって正しく捕捉されることにより動原体微小管が安定化することが、Kif18Aのプラス端集積の引き金となります。前中期初期の動原体微小管や、中期でも動原体微小管以外の紡錘体微小管のプラス端は比較的動的であるため、Kif18Aの集積が引き起こされません。しかし、微小管脱重合阻害剤であるTaxolを添加すると5分以内に細胞内のすべての微小管の先端にKif18Aが集積する様子が観察できます(movieへ)。
微小管プラス端動態の制御因子であるKif18Aの局在が、微小管プラス端動態によって制御されているというこの相互制御機構が、染色体の整列やオシレーション運動という繊細な運動を可能にする基盤となっていると考えられます
また、Taxol添加による微小管の安定化が、分裂期細胞内では微小管の異常伸長ではなく複数のアスター形成を引き起こす理由は長く謎でしたが、私たちの研究によりKif18Aの活性がこの現象に必要であることがわかりました(Masuda N., and Shimodaira T., et al., Cell Struct. Funct., 2, 261-267, 2011)。
現在、Kif18Aのtail領域がKif18Aの局在や活性に対してどのような役割を担っているかについて、解析を進めています。