研究紹介
3. 植物SRY遺伝子:雄を決定する遺伝子と性染色体
ナデシコ科の雌雄異株植物ヒロハノマンテマ(Silene latifolia)の雌雄は性染色体によって決定されています。多くの哺乳類と同じように、Y染色体をもつ個体が雄(♂)になります。Y染色体には雌蕊(しずい)を抑制する領域、雄蕊(ゆうずい)を促進する領域、葯(やく)を成育させる領域に加えて、X染色体と対合する領域(PAR)があり、そのどこかに雄を決める遺伝子、植物のSRY遺伝子があるはずです。ところが、Y染色体をもたない雌(♀)に黒穂菌(くろぼきん)が感染すると、雌花に雄蕊ができて雄(♂)化してしまいます。植物の雌雄決定カスケードでY染色体が果たす役割の一部を黒穂菌が肩代わりしているように見えます。黒穂菌は植物の雌雄がどうやって決まるか、植物SRY遺伝子を頂点とする雌雄を決定するカスケードのスイッチが何であるか知っているかのようです。
- 雌雄異種植物
- Silene latifolia (ヒロハノマンテマ)
- ヒロハノマンテマの性染色体
- マンテマ属の進化
- 植物性染色体の多様性
- クロボ菌が感染したヒロハノマンテマ
- 黒穂菌によって性転換させられる雌(♀)
- 健常雌(♀)と黒穂菌感染雌(♀)の花原基
- 黒穂菌感染雌(♀)のSLM2遺伝子の発現パターン
- ヒロハノマンテマの雄蕊発達制御メカニズム
3-1. 雌雄異種植物

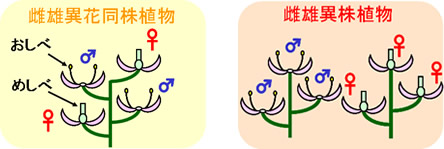
被子植物の生殖システムは多様です。被子植物の70%はシロイヌナズナに代表される両性花植物です。残り30%が雌雄異花同株あるいは雌雄異株植物で、トウモロコシ、キュウリ、カボチャに加え、ホウレンソウ、アスパラガス、ホップなど重要な農作物も多く含まれています。
スーパーで野菜と果物それにビールを買ってきました。ビールはホップのつもりです。さて、上の写真のなかで雌雄異株植物はどれとどれでしょう? 下は居酒屋メニューによくある、ホウレンソウのおひたし、アスパラのベーコン巻き、アスパラコーンですが、さて雌雄異株植物でないものはどれでしょう?

上の写真で雌雄異株植物は、ホウレンソウ、アスパラガス、キウイフルーツ、それにビールのホップが雌雄異株植物で、ビールにはホップの雌花 (♀)を使います。居酒屋メニューでは、コーン(トウモロコシ)だけが雌雄異株植物ではありません。トウモロコシやキュウリ、カボチャは、雌雄異花同株で、同じ株に雌花と雄花がつきます。
3-2. Silene latifolia (ヒロハノマンテマ)

ナデシコ科雌雄異株植物ヒロハノマンテマの花は恐らく雌雄異株植物のなかで最も清楚で美しい花に1つです。花冠から白い5本の花柱をのぞかせた雌花は、豊満な子房をくるむ合着した萼の黄緑が清楚ではあるが豊穣な印象を与えているのかも知れません。
北隆館の『牧野新日本植物図鑑』によると、ヨーロッパ、北アフリカ、北西アジアの原産で、多年草の草本とあって、5~6月から9月にわたって、枝分かれしてまばらな円錐花穂を出し、短い柄の上に白花をつけ、夕方開いて香気を出すとあります。マンテマは夜の花なのです。
ところで・・・「マンテマ」という不思議な和名の由来は、牧野の植物図鑑には、「海外から渡って来た当時の呼び名のマンテマンの略されたもので、このマンテマンは多分にAgrostemma(ムギセンノウ)という属名が転訛したものではないかと想像する。」と書かれています。因みに、「センノウ」は、嵯峨の仙翁寺にあったナデシコというような意味です。
参考文献
- 牧野富太郎「牧野新日本植物図鑑」北隆館、1060p (1961).
3-3. ヒロハノマンテマの性染色体
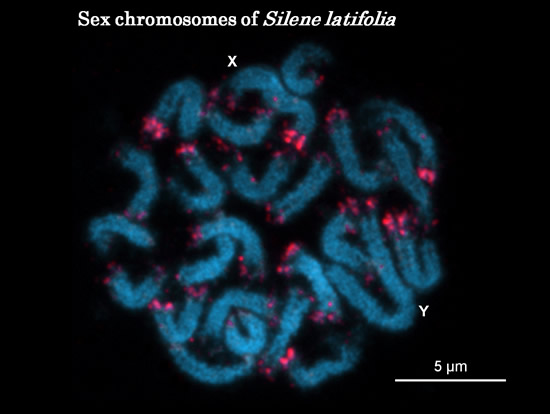
ヒロハノマンテマはXY型の性染色体をもちます。核型は22A+XYで、Y染色体が最も大きく、およそ570MbのDNAを含みます。Y染色体が中部動原体型であるのに対し、X染色体は次中部動原体型で、短腕(p)と長腕(q)との長さが大きく異なります。
植物でも性染色体には、組換えを起こさない領域と組換えを起こす領域(Pseudo Autosomal Reagions: PAR)があり、組換えを起こさない領域にはY染色体固有のDNA配列が蓄積し,そこに性決定因子が存在すると考えられています。雌雄異種植物で性染色体をもつヒロハノマンテマは植物の雌雄性、特に雄(♂)を決める遺伝子(SRY)の研究には打って付けの材料ということになります。
ヒロハノマンテマの染色体は大きくて顕微鏡で観察しやすいので、70年も前から性染色体が観察され、Y染色体に雄(♂)を決定する領域があることなども明らかにされています。植物のFISH法が確立される90年代になると、ヒロハノマンテマでも様々なプローブが単離され性染色体の構造解析がなされるようになります。ヒロハノマンテマで最初に単離されたFISHプローブは、フローサイトメトリーで分画したX染色体からDOP-PCRで増やしたX43.1だが (Buzeck et al. 1997)、Y染色体からはレーザーマミピュレーション技術で単離したRMY1 (Matsunaga et al. 1999)、反復単位にSacIやKpnIの認識配列もったサテライトDNA(Garido-Ramos et al.1999, Kazama and Kawano 2008)なども単離されています。これらのどの配列をプローブに用いてFISHをしても、染色体の末端にシグナルが得られます。興味深いのは、性染色体で、X染色体には両端にシグナルが得られたが、Y染色体でシグナルが検出されたのは片腕だけです。この非対称性は、XY性染色体の起源を考える上で重要な手がかりになりそうです。
因みに、DNAの縦列型繰り返し配列のうち、繰返し単位1-5bpをマイクロサテライト、繰返し単位6-40bpをミニサテライト、繰返し単位40bp以上をサテライトDNAと呼びます。私たちが見つけたKpnI は繰返し単位が313もあります。
参考文献
- Kazama, Y., Sugiyama, R., Matsunaga, S., Shibata, F., Uchida, W., Hizume, M., Kawano, S. (2003) Organization of the KpnI family of chromosomal distal-end satellite DNAs inSilene latifolia. J. Plant Res. 116:317-326.
- Kazama, Y., Sugiyama, R., Suto, Y., Uchida, W., Kawano, S.: The clustering of four subfamilies of satellite DNA at individual chromosome ends in Silene latifolia. Genome 49:520-530 (2006).
- Kazama, Y., Kawano, S.: Technical note-Detection of pseudo autosomal region in theSilene latifolia Y chromosome by FISH analysis of chromosomal distal end satellite DNAs. Cytologia 73, 1- 2 (2008).
- Ishii, K., Amanai, Y., Kazama, Y., Ikeda, M., Kamada, H., and Kawano, S.: Analysis of BAC clones containing homologous sequences on the end of the Xq arm and on chromosome 7 in the dioecious plant Silene latifolia. Genome, 53, 311-320 (2010).
3-4. マンテマ属の進化

種子植物全体を見渡すと、雌雄異株は目や科のレベルで広く植物界に分付しているように見えるが、それは単一の祖先から進化したようには決して見えません。その起源は1つではなく、雌雄異株は科や目のレベルでそれぞれ別々に出現したと考えられます。
雌雄異株植物の花の発達過程を観察すると、花器官の発達のごく初期には雄蕊と雌蕊の原基を両方もち合わせている種も多いことがわかります。これらの研究からも、ダーウィンが考えたように、両性花タイプが祖先型であり、両性花植物の雄蕊あるいは雌蕊の発達が別々に抑制されることによって雌雄異株植物が生まれたことが分かります。植物の性染色体もおなじで、雌雄異株の誕生とともに、科や門のレベルでそれぞれ独立に生じたと考えられています。
マンテマ属の系統に関してはDesfeuxら(1996)が作成した分子系統樹があります。上の図はそれにYildizらが撮った染色体の写真を貼り付けたものですが、ナデシコ科のなかでも雌雄異株と性染色体がそれぞれ独立に生じて進化してきたのが分かります。この系統樹からヒロハノマンテマは、800~2,400万年前に雌雄同株から雌性両全異株を経て進化し、そのときXY型の性染色体も獲得したとされています。哺乳類の性染色体の起源は約3億年前とされているので、ヒロハノマンテマの性染色体は極めて新しいことになります。古いものの起源より、新しいものの起源のほうが詳しく正確に調べられるでしょう。植物の性染色体研究の進捗が期待されます。
参考文献
- Desfeux, C, Maurice, S, Henry, JP, Lejeune, B, Gouyon, PH: Evolution of reproductive systems in the genus Silene. Proc R Soc B, 263: 409–414 (1996).
- Yildiz, K. and Çirpici, A.. 1996. Karyological studies on species of Silene L. northwest Anatolia. Turk. J. Bot. 20: 73–82.
3-5. 植物性染色体の多様性
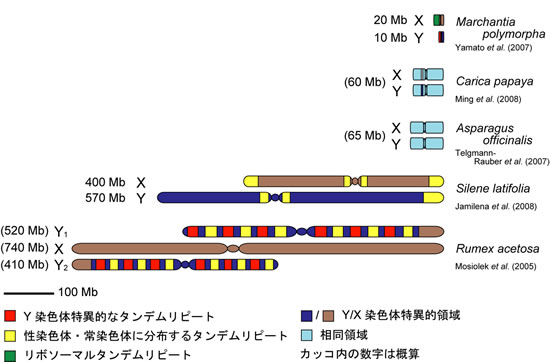
植物性染色体の出現と異形化の過程は様々な種の性染色体を調査することで類推できるでしょう。最も形が似ていて、原始的だと考えられる性染色体は、アスパラガス(Asparagus officinalis var.altilis.)やホウレンソウ(Spinacia oleracea)で観察されます。それらは、XY間で形状が同じで、Y(あるいはX)に固有の染色体領域は極めて小さく、組換え抑制領域はほとんど観察されません。パパイア(Carica papaya)の性染色体はXYの形状は顕微鏡的には同じに見えますが、そのうち10%に相当する領域で組換えが抑制されY(あるいはX)に固有の配列をもつことが分かっています。
X染色体とY染色体の間で最も形が異なっているのは、ヒロハノマンテマ(Silene latifolia)やスイバ(Rumex acetosa)の性染色体です。ヒロハノマンテマのY染色体はX染色体の約1.5倍の大きさで、そのうちの90%の領域で組換えが抑制されています。スイバでは、Y染色体が2本存在し、2本のY染色体はX染色体よりもやや小さい。その雌雄性は、ショウジョウバエのように、X染色体と常染色体の量比によって決定されています。X/Aが1以上ならばメスに、0.5以下ならばオスになります。スイバの性染色体でも大部分が組換え抑制領域です。ヒロハノマンテマとスイバで、最初に性染色体が生じたのは、前者で約2,000万年前、後者で約1,500万年前と推定されています。これらは恐竜が滅びたずっと後の新生代第三期にあたります。哺乳類の性染色体が誕生したのは3億年前の石炭紀であることと比較すると、植物性染色体は極めて新しいことになります。植物性染色体は組換え抑制とその領域拡大という性染色体進化の根幹を理解するための有力な研究材料になると期待されています。
参考文献
- 風間裕介・河野重行:ヒロハノマンテマの性染色体構造と性発現.Plant Morphology 21, 71-77 (2009)
- 風間裕介・河野重行:雌雄異株性―性染色体ならびに雄蕊(♂)と雌蕊(♀)の選択的発達制御.遺伝,63(3),42-47 (2009).
- 杉山立志・河野重行:両性具有の芳醇―パパイヤの性染色体―.遺伝 58(3), pp. 17-19 (2004).
3-6. クロボ菌が感染したヒロハノマンテマ

ヒロハノマンテマのY染色体上には、雄蕊(♂)の初期発達促進因子(SPF, Stamen promoting factor)や、雄性決定遺伝子(SRY)の存在が予測されています。XY染色体で遺伝的に決定されているヒロハノマンテマの雌雄性は植物ホルモンや光・温度などの環境要因には影響されません。SPFはY染色体のみにコードされているので、X染色体しかもたない雌(♀)には雄蕊(♂)を伸長させる能力はありません。
一方、黒穂菌(Microbotryum violaceum)が感染すると、雌花(♀)でも雄蕊(♂)が発達して両性花のようになります。これは、リンネ(1707~1778)の頃から知られていることで、黒穂菌は葯に胞子を形成するから、雌花(♀)で雄蕊(♂)を誘導するのは、黒穂菌の巧みな生存戦略の一つと考えられます。黒穂菌はY染色体の代わりをしていることになります。下の写真は左の小さいのが黒穂菌の胞子で右がヒロハノマンテマの花粉です。さらに左の人物がリンネです。

本郷キャンパスにある理学部2号館には小さな温室があります。その温室の前庭でヒロハノマンテマを栽培していたところ、偶然、黒穂菌が感染したヒロハノマンテマを見つけました。ヒロハノマンテマは日本には自生していないので、ヒロハノマンテマに感染したこの黒穂菌はどこからやって来たのか不思議です。右側の写真がそれで雄株に感染して葯のなかで黒穂菌の胞子を作ります。右は感染していない野生株の雌雄と感染株の雌雄です。雄(♂)も雌(♀)も花冠から黒い胞子がのぞいています。
3-7. 黒穂菌によって性転換させられる雌(♀)

雄蕊(♂)が伸張し葯が成熟するステージ11~12の頃の花を花弁を撮って観察すると、雌花(♀)に両性花のように雄蕊(♂)が生えてくるのがよく分かります。雄蕊(♂)とのトレードオフのせいか子房の発達が抑えられているように見えます。
左側が健常株の雌雄、右が感染株の雌雄、上が雄(♂)、下が雌(♀)です。左下を見ると雌花(♀)に生えてきた雄蕊(♂)の葯にも黒穂菌の胞子が作られ黒くなっているのが分かります。
黒穂菌は葯の中に胞子を作ることで訪花昆虫の助けをかりて伝播します。雌雄異種植物の場合、雌花(♀)は雄蕊(♂)を作らないので、何か工夫をしない限り伝播できないことになります。その工夫が性転換で雌(♀)に雄蕊(♂)を作らせることで伝播を達成するのです。
3-8. 健常雌(♀)と黒穂菌感染雌(♀)の花原基
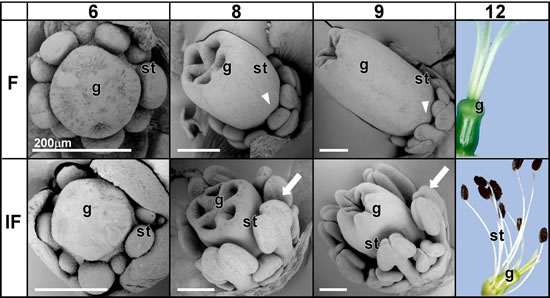
雌蕊(♀)や雄蕊(♂)の原基が発達するステージ6-9を走査電顕で観察しました。また、ステージ12になると、黒穂菌は雄蕊(♂)の葯内に胞子を作るので,感染した葯は黒く見えます(IFのステージ12)。ステージ6では、雌(♀)であっても雄蕊(♂)と雌蕊(♀)の原基の両方をもちます。健常株では、雌蕊(♀)が発達して5枚の心皮となりますが、雄蕊(♂)は抑制されたままです。一方,雌花(♀)ではあっても、黒穂菌感が感染していると、雄蕊(♂)は抑制されません。雌蕊(♀)も発達するので雄蕊(♂)が発達して雌花(♀)は両性花のようになります。
参考文献
- Uchida, W., Matsunaga, S., Sugiyama, R., Kazama, Y. and Kawano, S.: Morphological development of anthers induced by the dimorphic smut fungus Microbotryum violaceum in female flowers of the dioecious plant Silene latifolia. Planta 218: 240-248 (2003) .
- Uchida, W., Matsunaga, S. and Kawano, S.: Ultrastructural analysis of the behavior of the dimorphic fungus Microbotryum violaceum in fungus-induced anthers of femaleSilene latifolia flowers. Protoplasma 226: 207-216 (2005) .
3-9. 黒穂菌感染雌(♀)のSLM2遺伝子の発現パターン
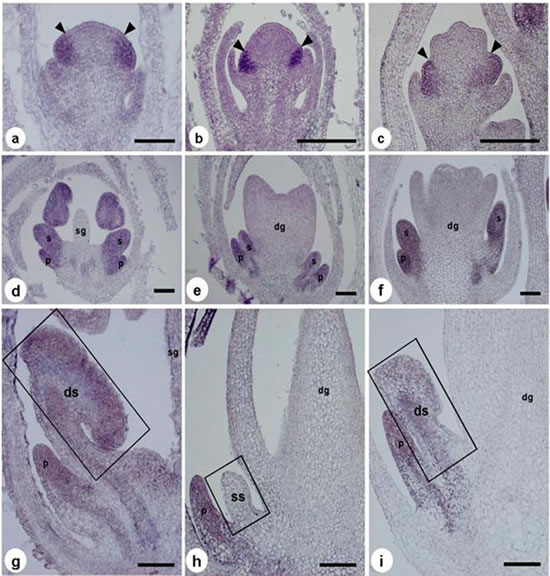
Y染色体の雄蕊(♂)の初期発達促進領域にコードされているのは、雄蕊(♂)の発達制御に関わるMADSボックス遺伝子やそれを制御する遺伝子だろうと考えられますが、黒穂菌はそれらの遺伝子の発現を撹乱している可能性があります。
ABCモデルでは、雄蕊(♂)と花弁のアイデンティティを決定をするB機能遺伝子APETALA3 (AP3)とPISTILLATA (PI) は雄蕊(♂)の成熟にも不可欠であることがわかっています。B機能遺伝子SLM2の発現をin situ ハイブリダイゼーションで調べました。SLM2はシロイヌナズナのPIのホモログです。ステージ3では、SLM2は、雄(♂)、雌(♀)、感染雌(♀)ともに、予定花弁領域と予定雄蕊(♂)領域で発現しています。ステージ8では、SLM2の発現は雌(♀)の雄蕊原基で消失しますが、感染雌(♀)と雄(♂)の雄蕊原基では発現が続いています。ステージ10では感染雌(♀)と雄(♂)で雄蕊(♂)にシグナルがみられます。
上の写真は、ヒロハノマンテマの健常雄(♂),健常雌(♀)と黒穂菌感染雌(♀)のSLM2遺伝子の発現パターンを示すin situ ハイブリダイゼーションです。a-cはステージ5と6、d-fはステージ7、g-iはステージ8で、それぞれSLM2をプローブにしたin situハイブリダイゼーション,ステージ5-7ではほとんど同じ発現パターンですが、ステージ8になると健常雌(♀)の抑制雄蕊(♂)では発現が抑制されますが、黒穂菌感染雌(♀)では発現が続行します。
参考文献
- Kazama, Y., Koizumi, A., Uchida, W., Ageez, A. and Kawano, S.: Expression of the floral B-function gene /SLM2 /in female flowers of Silene latifolia infected with the smut fungus Microbotryum violaceum. Plant Cell Physiol. 46, 806-811 (2005).
3-10. ヒロハノマンテマの雄蕊発達制御メカニズム
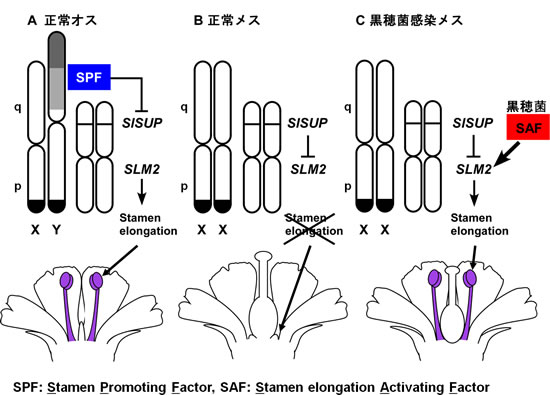
雌花(♀)と黒穂菌感染雌花(♀)でSlSUPの発現を調べました。雌花(♀)では、雄蕊の発達が止まるステージにSlSUPの発現量が上昇しますが、黒穂菌感染雌花(♀)では、逆にSlSUPの発現量が低下していました。ヒロハノマンテマの雌花(♀)では、SlSUPのはたらきにより積極的に雄蕊フュ達が抑制されていると考えられます。Y染色体をもつ雄(♂)ではSlSUPは発現しません。Y染色体の「雄蕊の発達を促進する遺伝子」がSlSUPの発現を抑制しているらしく、雄(♂)で雄蕊(♂)が伸長するのはこのためであろう。メスには「雄蕊の発達を促進する遺伝子」が無いため、SlSUPが発現し雄蕊(♂)の伸長が抑制されるということになるのでしょう。黒穂菌感染雌花(♀)でも、SlSUP発現と雄蕊(♂)伸長抑制の間には明らかな相関が見られたことから、黒穂菌はSlSUP発現を抑制することで雄蕊(♂)の発達を促進しているのではないかと考えられます。
植物はそのほとんどが両性花をつけるので、雌雄性への関心が動物に比べて稀薄なように思われます。雌雄異株植物には農業や他の産業に有用なものも多いのにもかかわらず、研究者人口は思いのほか少ないように思われます。ヒロハノマンテマなどでは遺伝子操作技術の開発も有]ですし、ゲノム解読技術進歩には目覚しいものがあり、パパイアのほかにもポプラでは全ゲノムが解読され性染色体の構造も解析が進んでいます。アスパラガスやホウレンソウでも組換え抑制領域が同定されています。Sry遺伝子は勿論、組換え抑制領域のDNA配列が解読されるのも間近でしょう。
参考文献
- 風間裕介・河野重行:雌雄異株性―性染色体ならびに雄蕊(♂)と雌蕊(♀)の選択的発達制御.遺伝,63(3),42-47 (2009).idennsi
続きます。お楽しみに・・