搶嫗戝妛 惗暔惗嶻岺妛尋媶僙儞僞乕
娐嫬曐慡岺妛尋媶幒
尋媶撪梕 - 怉暔婡擻偺夝愅
1丗怉暔偺昦奞掞峈惈敪尰婡峔偺夝柧
変乆偑儌僨儖宯偲偟偰梡偄偰偄傞僀僱偵偍偄偰偼丄昦尨嬠桼棃偺惉暘偱偁傞僄儕僔僞乕偑嵶朎枌偵懚嵼偡傞庴梕懱偲寢崌偡傞偙偲偵傛傝昦尨嬠姶愼偑擣幆偝傟偰偄傞丅偦偟偰丄偦傟偑堷偒嬥偲側偭偰丄僕儍僗儌儞巁側偳偺擇師僔僌僫儖丄僄儕僔僞乕丒僕儍僗儌儞巁墳摎惈偺揮幨場巕摍傪夘偟偨僔僌僫儖揱払婡峔偵傛傝峈嬠惈僞儞僷僋幙偺惗惉傗僼傽僀僩傾儗僉僔儞偲憤徧偝傟傞峈嬠惈擇師戙幱嶻暔偺惗嶻側偳偺條乆側杊屼墳摎偑桿摫偝傟偰偄傞(恾1-1)丅
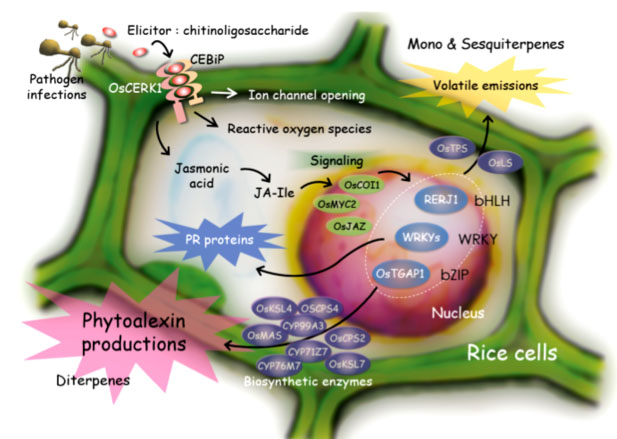
恾1-1. Defense mechanisms against pathogens in rice cells.
変乆偼丄忋婰偺傛偆側僄儕僔僞乕偺揱払婡峔傪夝柧偟丄偦偺惉壥傪娐嫬曐慡宆擾嬈媄弍偺奐敪乮昦奞掞峈惈桿摫宆擾栻扵嶕朄偺妋棫丄昦奞懴惈僀僱偺嶌弌側偳乯偵墳梡偡傞偨傔丄埲壓偺傛偆側尋媶傪峴偭偰偄傞丅
(1)僀僱偵偍偄偰偼15庬椶偺壔崌暔偑僼傽僀僩傾儗僉僔儞偲偟偰摨掕偝傟偰偄傞偑丄偦偺偆偪偺14庬椶偼僕僥儖儁儞宆壔崌暔偱偁傞丅変乆偼丄僕僥儖儁儞宆僼傽僀僩傾儗僉僔儞惗崌惉偵娭梌偡傞僕僥儖儁儞娐壔峺慺堚揱巕傪偡傋偰扨棧摨掕偡傞偲偲傕偵乮嶳宍戝妛偲偺嫟摨尋媶乯丄偦偺忋棳丄壓棳偺惗崌惉堚揱巕偵偮偄偰傕夝愅傪恑傔丄偦偺惗崌惉宱楬偺慡梕偑柧傜偐偵側傝偮偮偁傞乮恾1-2乯丅
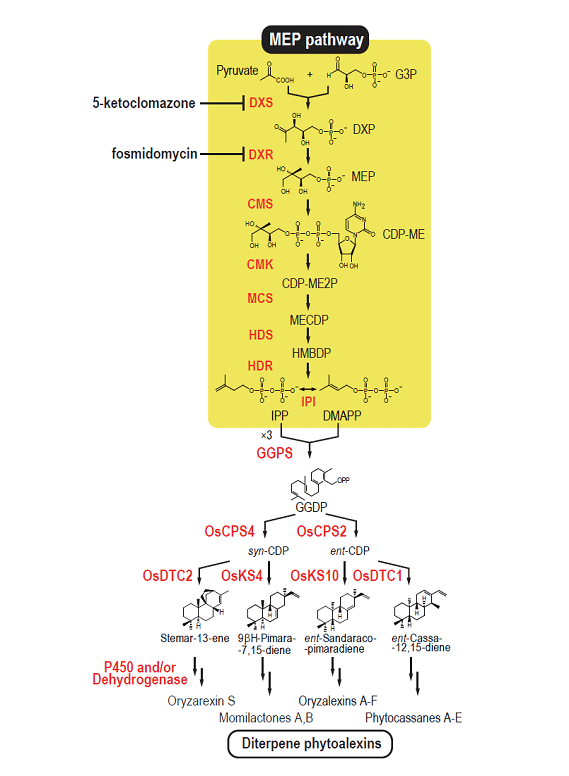
恾1-2. Biosynthetic pathways for diterpenoid phytoalexins in rice.
傑偨丄僼傽僀僩傾儗僉僔儞惗崌惉偺惂屼婡峔偵偮偄偰傕夝愅傪峴偭偰偍傝丄僼傽僀僩傾儗僉僔儞惗崌惉峺慺堚揱巕偑丄bZIP宆傗WRKY宆揮幨場巕偺惂屼壓偵偁傞偙偲丄庡梫僼傽僀僩傾儗僉僔儞偱偁傞儌儈儔僋僩儞椶丄僼傽僀僩僇僒儞椶偺惗崌惉堚揱巕偑僋儔僗僞乕傪宍惉偟偰偄傞偙偲側偳偑帵偝傟偰偄傞丅
(2)僉僠儞僆儕僑摐側偳偺僄儕僔僞乕傪張棟偟偨僀僱偺攟梴嵶朎偐傜扨棧偟偨丄2庬偺僄儕僔僞乕墳摎惈偺 WRKY宆揮幨場巕 (OsWRKY53丄OsWRKY71) 偍傛傃1庬偺bHLH宆揮幨場巕RERJ1偵偮偄偰夝愅傪恑傔丄OsWRKY53偵娭偟偰偼丄摉奩堚揱巕偺夁忚敪尰懱傪梡偄偨儅僀僋儘傾儗僀夝愅丄媦傃偄傕偪昦嬠愙庬幚尡偐傜丄僀僱偺昦奞掞峈惈偵娭梌偡傞揮幨場巕偱偁傞偙偲傪柧傜偐偵偟偨乮恾1-3乯丅
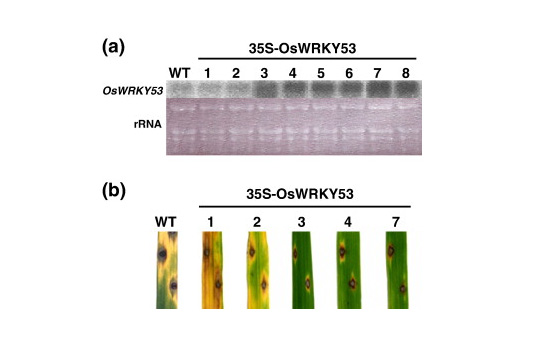
恾1-3.Effects of overexpression of OsWRKY53 on resistance to M. grisea race 007 in rice plants. (a) Overexpression of OsWRKY53 in transgenic rice plants. Northern analysis was performed using total RNA (10μg) prepared from transgenic rice plants. Ethidium bromide staining of rRNA confirmed equal loading of total RNA in each lane. The numbers at the top of the gels indicate independent transgenic rice cells. VC, vector control. (b) Responses of leaf blades of 35S-OsWRKY53 transgenic rice plants to infection by the compatible M. grisea race 007. Lesions were observed 1 week after infection.
堦曽丄OsWRKY71偵娭偟偰傕摨條偵丄摉奩堚揱巕偺夁忚敪尰懱傪梡偄偨儅僀僋儘傾儗僀夝愅偵傛傝丄僀僱偺昦奞掞峈惈偵娭梌偡傞揮幨場巕偱偁傞偙偲傪柧傜偐偵偟偨丅傑偨丄RERJ1偼僀僱怉暔懱偵偍偄偰傕丄昦奞掞峈惈敪尰偵廳梫側栶妱傪壥偨偡怉暔儂儖儌儞偺僕儍僗儌儞巁傗丄偦偺惗嶻桿摫傪懀偡彎奞張棟偵傛偭偰丄塻晀偱堦夁揑側敪尰傪帵偟偨丅尰嵼丄偙傟傜偺揮幨場巕傪拞怱偲偟偨堚揱巕僱僢僩儚乕僋傪夝柧偡傞偙偲傪栚揑偲偟偰丄摉奩揮幨場巕偺昗揑堚揱巕孮偺僗僋儕乕僯儞僌丄媦傃摉奩揮幨場巕堚揱巕偺敪尰惂屼婡峔偺夝愅傪峴偭偰偄傞丅
(3)僀僱偺杊屼墳摎偵娭傢傞僕儍僗儌儞巁偺婡擻偺堦偮偲偟偰12-oxophytodienoic acid reductase 1 (OsOPR1)偺敪尰桿摫偑嫇偘傜傟傞丅OsOPR1偼僕儍僗儌儞巁惗崌惉峺慺偺堦偮偱偁傞壜擻惈偑峫偊傜傟偰偄偨偑丄僀僱偵偼彮側偔偲傕10庬偺儂儌儘僌偑懚嵼偟丄OsOPR7偲柦柤偟偨僷儔儘僌偑僕儍僗儌儞巁惗崌惉偵娭梌偡傞OPR偱偁傞偙偲偑幚徹偝傟偨(恾1-4)偙偲偐傜丄
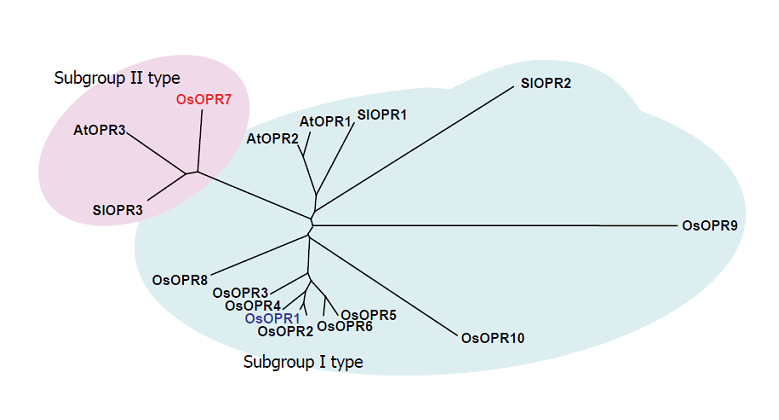
恾1-4.Phylogenetic relationship of OPRs between rice and other plants. The amino acid sequences deduced from OPR cDNA of rice (OsOPRs), tomato (SlOPRs), and Arabidopsis (AtOPRs) were compared using the program CLUSTALW. The phylogenetic tree was generated from the resulting alignment using the program TREEVIEW. Subgroup I and II type OPRs are indicated by shaded circles.
尰嵼偱偼丄OsOPR1偼僗僩儗僗帪偵惗惉偡傞兛丒兝晄朞榓働僩儞峔憿傪桳偡傞撆惈暔幙偺夝撆偵娭梌偡傞壜擻惈偑峫偊傜傟偰偄傞丅側偍丄OsOPR1偺敪尰惂屼偵偼摉奩堚揱巕偺僾儘儌乕僞乕夝愅偵婎偯偄偰bZIP宆揮幨場巕偺娭梌偑帵嵈偝傟偨丅
2丗怉暔偺僥儖儁僲僀僪惗崌惉弶婜抜奒偵娭偡傞尋媶
怉暔偼旕忢偵懡偔偺擇師戙幱嶻暔傪惗嶻偡傞偑丄偦偺拞偱傕僥儖儁僲僀僪壔崌暔偵暘椶偝傟傞暔幙偼丄帺慠奅偵2枩庬埲忋懚嵼偡傞偙偲偑抦傜傟偰偄傞丅忋婰僥乕儅偱弎傋偨僼傽僀僩傾儗僉僔儞傕丄怉暔偺昦奞掞峈惈敪尰偺嵺偵桿摫揑偵惗嶻偝傟傞扽慺悢20偺僕僥儖儁僲僀僪壔崌暔偺1偮偱偁傞丅
僼傽僀僩傾儗僉僔儞惗惉偵帄傞惗崌惉宱楬偼丄怉暔偵偍偄偰偼嵶朎幙偵懚嵼偡傞儊僶儘儞巁(MVA)宱楬偲僾儔僗僠僪偵懚嵼偡傞儊僠儖僄儕僗儕僩乕儖儕儞巁(MEP)宱楬偺擇偮偱峔惉偝傟偰偄傞偑丄僕僥儖儁儞宆偺僼傽僀僩傾儗僉僔儞崌惉偵偍偄偰偼MEP宱楬偑庡梫側宱楬偱偁傞偲梊憐偝傟偰偄傞丅
変乆偼丄僀僱偑奜奅偐傜偺僗僩儗僗偵墳偠偰僼傽僀僩傾儗僉僔儞傪惗崌惉偡傞帪偵偼丄MEP宱楬堚揱巕孮偺桿摫揑側妶惈壔偑堦惸偵婲偙傞偙偲傪尒偄偩偟偨丅MEP宱楬偼丄怉暔偺惗挿惗棟偵寚偐偣側偄僕儀儗儕儞丄傾僽僔僕儞巁丄僒僀僩僇僀僯儞側偳偺怉暔儂儖儌儞偺惗崌惉婲尮偱偁傞偙偲偐傜丄僀僱偼偙傟傜偺廳梫側怉暔儂儖儌儞偺惗崌惉偵塭嬁傪媦傏偡偙偲側偔丄昦奞掞峈偱妶桇偡傞僼傽僀僩傾儗僉僔儞傪廫暘惗嶻偡傞偨傔偵丄MEP宱楬堚揱巕偺嫤挷揑側敪尰桿摫傪峴側偆婡峔傪旛偊偰偄傞偲峫偊傜傟傞乮恾1-5乯丅
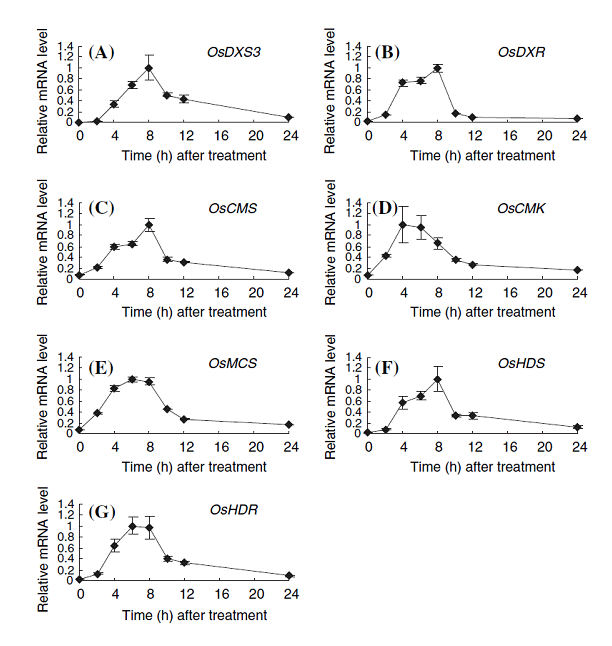
恾1-5. Expression profiles of possible genes in the MEP pathway in cultured-cells. The total RNAs used for expression profiling were isolated from rice cells exposed to 1 ppm chitin elicitor for the indicated times. Values indicate relative mRNA levels normalized to the expression of the UBQ gene, and the maximal value in each experiment with different primers was arbitrarily set to 1.0. A, OsDXS3; B, OsDXR; C, OsCMS; D, OsCMK; E, OsMCS; F, OsHDS; G, OsHDR. The results are the average of at least three independent experiments; bars indicate the standard deviation of the mean.
偝傜偵丄嵟嬤変乆偼丄僕僥儖儁儞宆僼傽僀僩傾儗僉僔儞偺惗嶻惂屼偺尞偲側傞揮幨場巕OsTGAP1傪敪尒偟偨丅偙偺揮幨場巕偼丄僀僱4斣愼怓懱忋偺儌儈儔僋僩儞惗崌惉峺慺堚揱巕僋儔僗僞乕偺敪尰惂屼傪峴偆偩偗偱側偔丄惗崌惉忋棳偺儊僠儖僄儕僗儕僩乕儖儕儞巁宱楬偺惂屼偵傕娭梌偡傞偙偲偐傜丄僕僥儖儁儞宆僼傽僀僩傾儗僉僔儞惗嶻偺儅僗僞乕惂屼場巕偱偁傞偲峫偊傜傟傞丅尰嵼偼丄偙偺OsTGAP1傪婲揰偲偡傞僀僱偺僥儖儁僲僀僪惗崌惉娭楢堚揱巕偺徻嵶側揮幨惂屼婡峔偵偮偄偰丄暘巕儗儀儖偱偺夝柧偵岦偗偨尋媶傪恑傔偰偄傞(恾1-6)丅
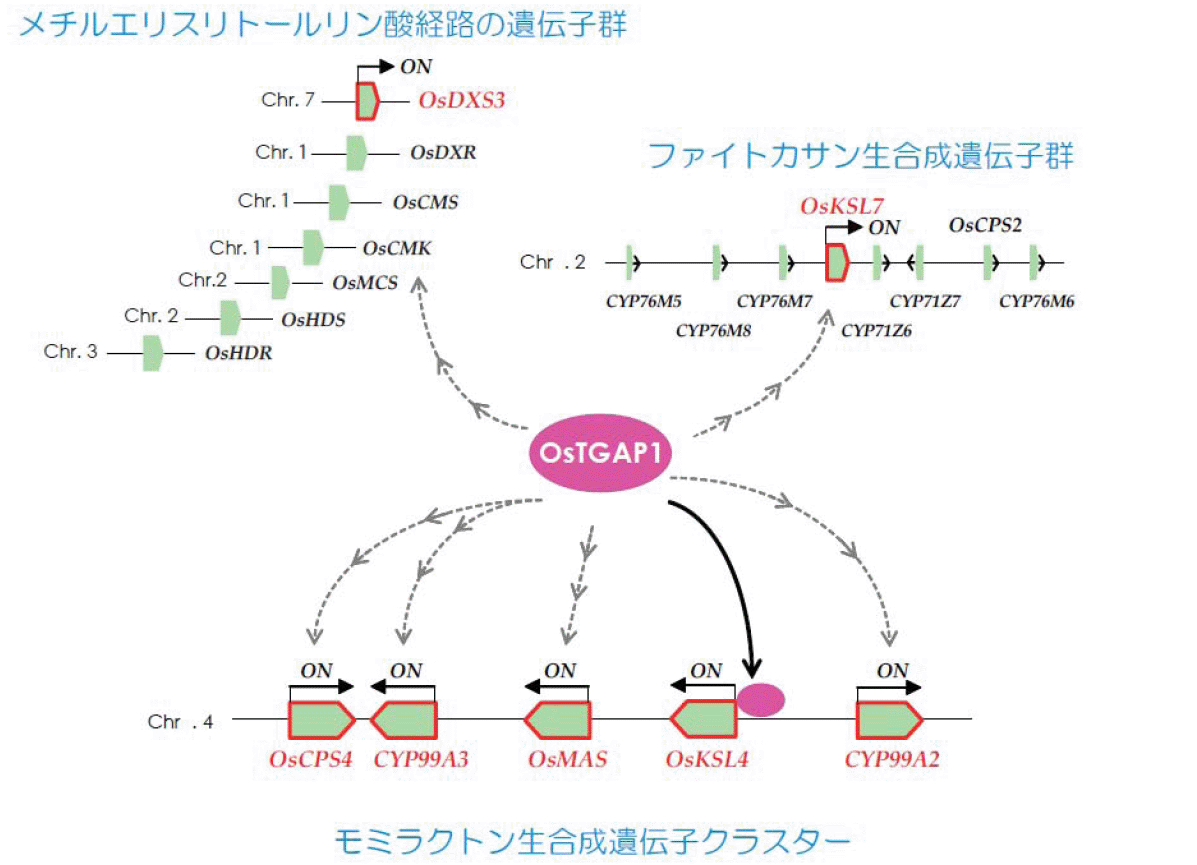
恾1-6. OsTGAP1 functions as a key regulator of the coordinated transcription of genes involved in inductive diterpenoid phytoalexin production in rice, and mainly exerts an essential role on expression of the clustered genes for momilactone biosynthesis.