「多細胞生物」麹菌における細胞間連絡のメカニズムの解析
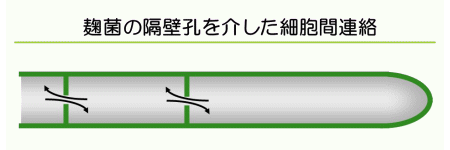
麹菌の菌糸は細長い細胞が連なってできていますが、これを仕切る隔壁には隔壁孔という小さな穴があいており、これを通じて隣り合う細胞は連絡しています。単細胞の酵母にはない、麹菌の多細胞のメカニズムについて解析しています。
麹菌は“多細胞”であり、その菌糸は多くの細長い細胞が連なってできています。これを仕切る隔壁には隔壁孔という小さな穴があいており、これを通じて隣り合う細胞は連絡しています(図1)。動物のギャップ結合や植物の原形質連絡のような、多細胞生物が恒常性を維持するための重要な細胞システムの一つであると考えられます。
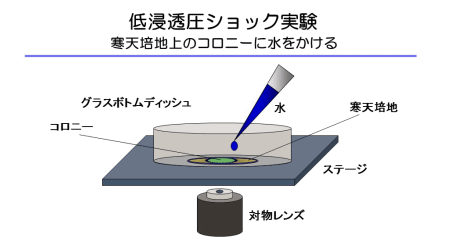
この隔壁孔を介した細胞間連絡はリスクをはらんでおり、ある細胞が損傷すると、隔壁孔を介してつながっている隣の細胞が巻き添えに遭う可能性があります。私たちは以前、麹菌を寒天培地に生育させ、コロニーに水をかけて低浸透圧ショックを与える実験を行い、菌糸先端から細胞内容物が噴き出して溶菌することを偶然発見しました(図2+動画1)。
動画1:低浸透圧ショックによる麹菌の溶菌
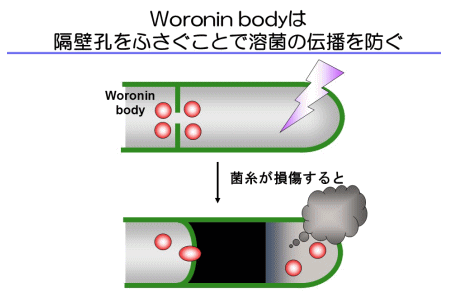
ところが、溶菌した先端細胞に隣接する2番目の細胞には溶菌は伝播せず、生き残りました。隣の細胞が巻き添えに遭うリスクを防ぐのが、Woronin bodyと呼ばれる糸状菌特異的なオルガネラです。Woronin bodyは隔壁の近傍に観察され、菌糸が損傷したときに隔壁孔をふさぐことで溶菌の伝播を防ぐ役割をもちます(図3)。私たちは、低浸透圧ショックによって麹菌が溶菌する性質を利用し、Woronin bodyが隔壁孔をふさぐことを、蛍光タンパク質を用いて初めて証明しました(動画2)。
動画2:隔壁孔をふさぐWoronin body
さらに、私たちはオランダのグループとの共同研究により、Woronin bodyが通常の生育条件でも隔壁孔をふさぐ現象を見いだしました。これは、溶菌時にのみ隔壁孔をふさぐという従来の定説を覆す発見でした。また、Woronin bodyを隔壁につなぎとめる巨大なタンパク質AoLAHの、天然変性領域におけるバネのような働きを明らかにしました(図4)。
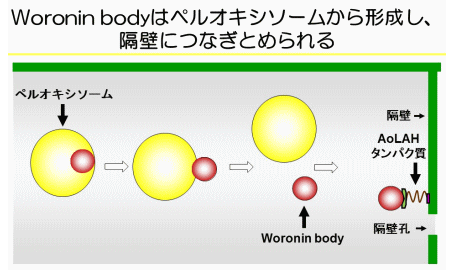
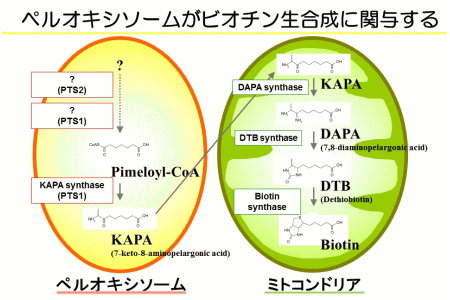
続いて私たちは、Woronin bodyがペルオキシソームという真核生物に普遍的に存在するオルガネラから分化して形成することを明らかにしました(図4)。この解析のなかで、麹菌のペルオキシソーム機能欠損株が、最少培地で生育できないことに気付き、偶然、ビタミンの一種であるビオチンの添加により、生育が回復することを突き止めました。一連の実験により、ペルオキシソームがビオチンの生合成に関与することを、世界で初めて発見することになりました(図5)。さらに、植物でも本発見を支持するデータを得て、ビオチンを生合成する真核生物に普遍的な現象を明らかにしました。
最近はWoronin body以外に、様々なストレスに応答して隔壁孔に蓄積するタンパク質、隔壁孔を囲う局在を示すシグナル分子など、隔壁孔における多細胞の恒常性維持に様々な因子が関与することを見いだしつつあります。私たちは麹菌をモデルとして、未だに多くが明らかになっていない、糸状菌の多細胞生物としての分子制御の総合的な解明に挑んでいます。
麹菌の細胞融合と不和合性の解析と有性生殖能の開発
麹菌では有性世代が見つかっておらず、以前は不完全菌として分類されてきました。様々な株の優良な形質を併せもつ株を育種するための交配が適用できないのが長年の課題となっています。
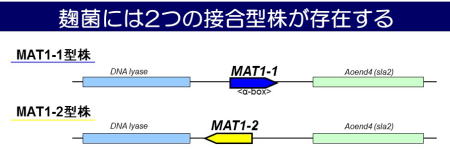
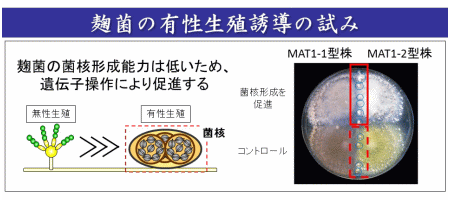
私たちはこれまでに、様々な用途の麹菌株を調べて2つの接合型(MAT1-1型, MAT1-2型)が存在することを明らかにしました(図1)。このことにより、麹菌がヘテロタリックな有性生殖を行う能力をもつ可能性が考えられました。現在は、有性生殖に関連する遺伝子を操作することにより、麹菌において有性生殖を誘導することを試みています(図2)。
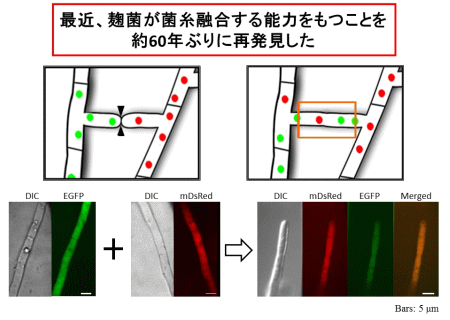
有性生殖の過程では、異なる接合型をもつ株どうしが細胞融合を行う必要があります。麹菌の細胞融合に関する研究は、1950年代に坂口謹一郎東京大学名誉教授らの報告がありましたが、以降は途絶えていました。私たちは、約60年ぶりに麹菌が細胞融合能をもつことを再発見しました(図3)。さらに、細胞融合に関与する新規因子を見いだすことで、麹菌の細胞融合能を向上させることを目指しています。
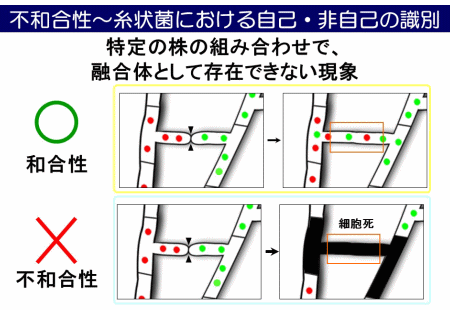
麹菌には日本酒・醤油・味噌など用途によって多種多様な株がありますが、私たちは最近、株の組み合わせによっては細胞融合体が存在できない「不和合性」という現象を発見しました。すなわち、同じ麹菌でありながらも、株間で自己・非自己を見分ける仕組みが存在することを明らかにしました。現在は、麹菌の不和合性の解消や有性生殖能力を高めるための研究を行っています。
麹菌におけるゲノム編集技術の利用による有用物質生産株の開発
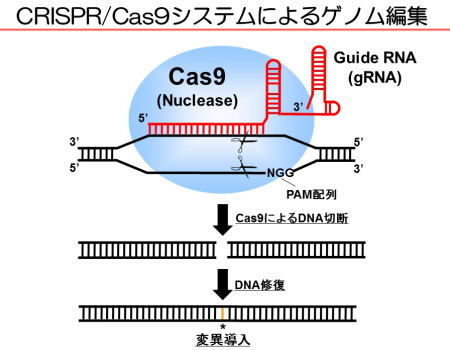
ゲノム編集は、部位特異的なDNA 切断酵素を用いて標的遺伝子を書き換える技術です。CRISPR/Cas9 システムは、ゲノム編集のなかでも最も安価で簡便な変異導入法であり(図1)、幅広い生物種に適用可能で、農業や医療分野への応用が期待されています。
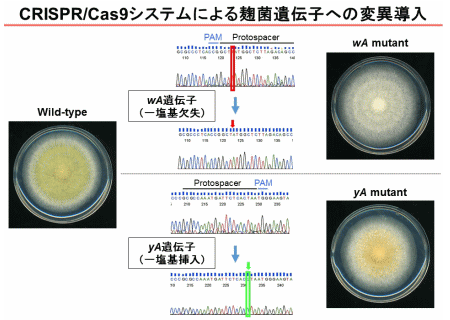
私たちは麹菌で初めて、CRISPR/Cas9システムを導入し、標的のゲノム配列を書きかえる技術を確立しました(図2)。麹菌実用株において従来は困難であった遺伝子操作の効率を飛躍的に向上することに成功しました。この技術を利用して、有用物質生産における麹菌の潜在能力を伸ばす研究を行っています。
麹菌が光に応答して分化を制御するメカニズムの解析
麹菌は、動物や植物と同じように光に応答する性質をもっています。最近私たちは、麹菌が光受容体をもつことを明らかにしましたが、この光応答のメカニズムを解明できれば、醸造や有用物質生産の現場で応用できるのではと期待されています。
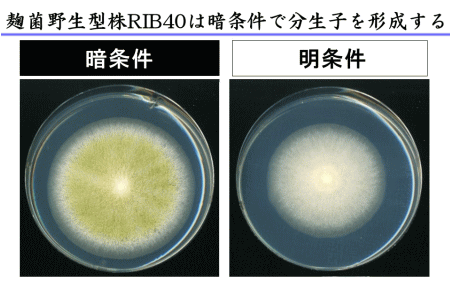
光は自然界に存在する環境因子の1つです。麹菌も、動物や植物と同じように光に応答する性質をもっていますが、これまでにあまり注目されてきませんでした。糸状菌において分生子形成は光によって促進されるのが一般的であるのに対して、麹菌の野生株であるRIB40株では逆に分生子形成が抑制されることが報告されています(図1)。
私たちは最近、発光ダイオードLEDを用いて、麹菌の光応答における波長の特異性を特定するとともに、光受容体に依存することを明らかにしています。麹菌の特異的な光応答のメカニズムを解明することにより、醸造や有用物質生産の現場での応用を期待して、研究を進めています。








