



植物の環境応答と耐性の基本的なしくみ
植物は、乾燥や高温、低温などの環境ストレスに応答し、さらに強いストレスにも耐えられるように自らを変化させる機構を進化させてきました。これは、自ら動いてストレスを回避することはできない植物のストレス応答の大きな特徴です。この機構は当研究室も含めた世界中の研究グループにより解析され、環境ストレスを受けた植物の細胞では、ストレスに応答したダイナミックな遺伝子発現変動により細胞内外の変化が引き起こされ、その結果、植物がそれぞれのストレスに耐えられるようになるという基本的な仕組みが明らかになりました(図1)。また、この遺伝子発現を制御することによって植物のストレス耐性を向上させることが可能であることも示されています。
一方で、植物がストレスをどのように感じているのか、つまりストレスの受容から植物体内、細胞内での伝達、遺伝子発現に至る初期過程がどうなっているのかについては不明な点が多く残されています。現在、私たちは、これらの過程の解明を目指した研究を行っています(1)(3)。また、作物を利用して、得られた知見の応用可能性を探る研究や新たな耐性機構を明らかにする研究を行っています(2)。さらにストレス応答を利用した作物の形質制御に関する研究も行っています(4)。
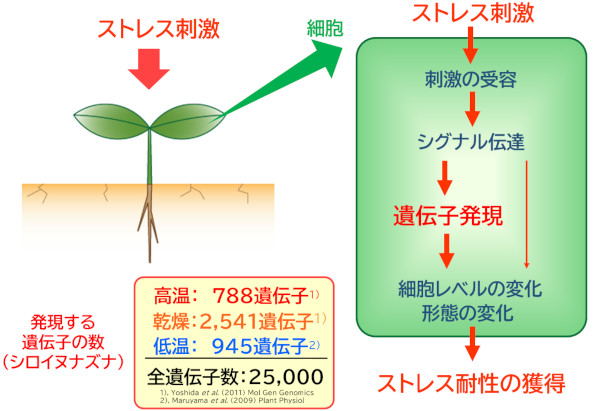
図1. 植物は環境ストレスを受けると、そのストレスに対する耐性が向上し、さらに強いストレスを受けた時にも耐えられるようになる。その鍵となるのがストレスに応答した遺伝子発現の変動で、これによる細胞内外の変化がストレス耐性を向上させる。
(1)環境ストレス応答の初期過程の解明
ストレスに応答した遺伝子発現の鍵となっているのが転写因子と呼ばれるタンパク質で、それぞれの転写因子は制御下にある複数の遺伝子の発現を同時に活性化することができます。また、図2のように、転写因子はストレスによって使い分けられており、転写因子はストレスシグナルの情報が集積するハブとして機能していることが分かります。これまでの研究でこれらの転写因子の活性を制御する機構が少しずつ明らかになってきました。現在は、環境条件によって転写因子の活性を制御するシグナル伝達系を明らかにし、植物がストレスをどのように感じているのか解明する研究を行っています。
現在、主要なテーマの一つとして解析しているのが、DREB2という転写因子を中心とした経路です(図3)。DREB2は高温と乾燥の両方のストレスで機能するというユニークな特徴を持っています。高温と乾燥は干ばつなどの状況で複合的に発生するなど互いに関連が深く、植物の生存や生殖に悪影響を及ぼすストレス条件です。この二つのストレスの両方に対応したDREB2経路はこれらのストレス応答に重要であると考えられ、実際にこの経路を人為的に活性化させることで、植物の高温や乾燥に対する耐性を向上させることができます。これまでにDREB2A経路の活性を制御するいくつかの機構を解明してきましたが、DREB2A経路がストレスによって直接活性化される仕組みは明らかになっていません。そこで、この活性化の仕組みやDREB2経路の正体(DREB2経路は細胞内のどのような状態を感知し、どのように変化させるのか)を明らかにすべく、解析に取り組んでいます。
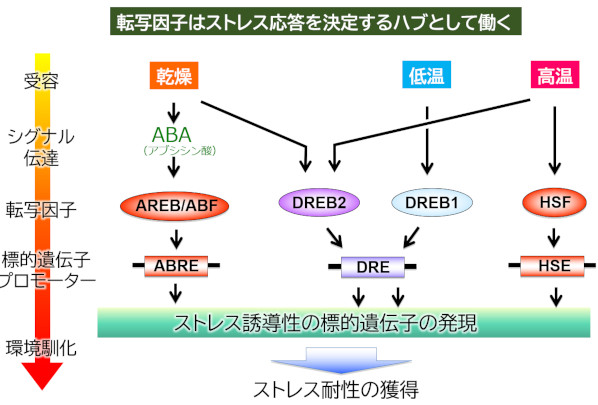
図2.ストレスに応答した遺伝子発現の鍵となっているのが転写因子と呼ばれるタンパク質群である。転写因子は標的遺伝子のプロモーターに結合して発現を制御するタンパク質で、一つの転写因子が複数の遺伝子を同時に制御することができる。
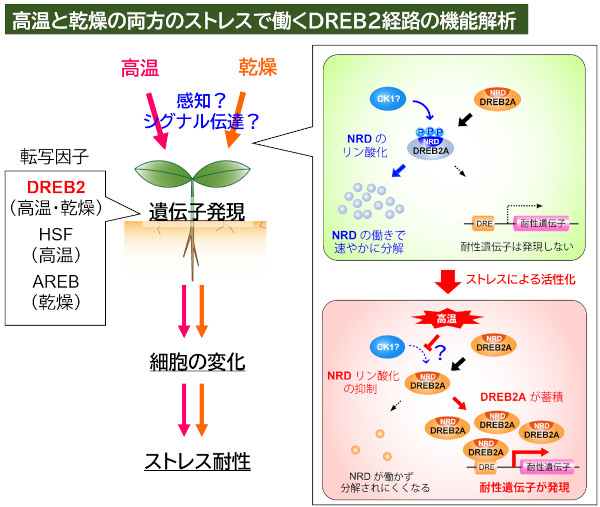
図3.DREB2経路は、高温と乾燥の両方のストレスへの応答で機能する転写因子で、これは、HSF、ABREがそれぞれ高温、乾燥へのストレス応答を主に担っているのに対して対照的な特徴である。DREB2経路の機能は通常時は抑制されており、ストレスの刺激が伝わると活性化されるが(Mizoi et al. 2019など)、活性化に至る上流機構は不明である。
(2)作物を用いたストレス応答研究と、作物のストレス耐性向上技術の開発
ストレス応答の研究から重要性が明らかになったタンパク質(転写因子やその活性制御にかかわるタンパク質、また下流の耐性機構にかかわるタンパク質)の遺伝子を作物に導入することで、ストレス耐性を向上させることができます。図4は、モデル植物のシロイヌナズナでDREB2Aと協調して高温耐性に働いていることが明らかになったタンパク質DPB3-1を、イネに導入した例です。イネにはDREB2Aと相同な転写因子であるOsDREB2Bが存在するためDPB3-1は高温応答を増強すると期待されましたが、実際にDPB3-1を導入したイネでは高温耐性が向上しました。また、図5の例では、ダイズのゲノム情報を利用して、DREB2Aと相同な機能を持つ転写因子、GmDREB2A;2を同定しました。
一方、トマトを用いた高温ストレス応答の研究から、トマトにおいてもシロイヌナズナと同様の複数の高温ストレス応答経路が存在していて、温度帯によってそれぞれの経路が使い分けられていることが明らかになりました。また高温耐性の品種間差が、HSFと呼ばれる転写因子によって制御される経路の頑健性と相関していることを見出しました(図6)。このことは、遺伝子発現を調べることでトマトの高温耐性度を推定できる可能性を示しています。
以上のように、シロイヌナズナと作物を用いた比較多様性研究によって、シロイヌナズナで得られた知見の一般性を確認するとともに、ストレス応答の進化についても明らかにしたいと考えています。また、それぞれの作物はモデル植物とは異なるさまざまな特性を持っています。そこで、作物を起点とした、新たなストレス応答機構の解明にも取り組んでいます。
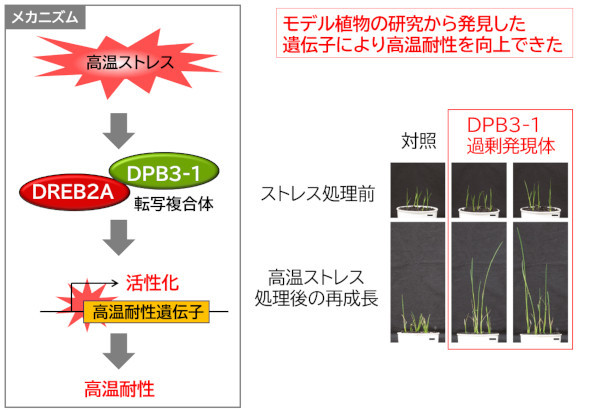
図4. モデル植物のシロイヌナズナの高温耐性の研究から明らかにした転写因子DPB3-1をイネに導入すると高温耐性が向上する(Sato et al. 2016)。

図5.ダイズにおいて乾燥ストレスに応答して働く転写因子GmDREB2A;2を発現するシロイヌナズナの形質転換体は、乾燥ストレスに強くなる(Mizoi et al. 2013)。
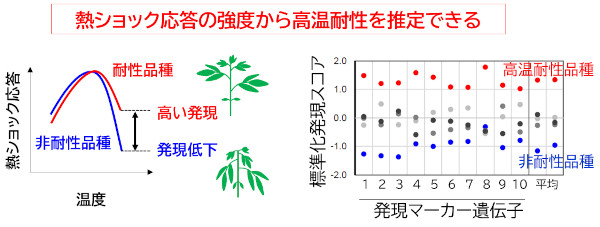
図6.高温耐性のトマト品種は、急激な熱ショック条件に対しても高温応答を維持できる能力を持っている。高温応答性遺伝子を発現マーカーとして利用することで、高温応答の強さを推定することができる(Mizoi et al. 2023)。
(3)環境ストレスにさらされた植物の生育制御機構の解明
環境ストレスにさらされた植物は、生育が抑制されることが知られています。この抑制は従来、光合成の減少や膨圧の低下によるものとされていましたが、近年の研究から、植物がストレス下で生育を積極的に抑制していることが明らかになってきました。図7はその一例を示しています。環境ストレス条件下で発現が抑制される遺伝子の解析から、ストレス下においては、草丈を伸ばす転写因子の機能が抑制されることが明らかになりました。また、ストレス応答に関わる転写因子の導入はしばしば生育も抑制します。
このように植物はストレスに対して、直接的な耐性機構だけではなく生育制御も含めた総合的な応答によって対応するよう進化してきました。自然界においては、環境条件が悪化したときに、更なる環境条件の悪化に備えて生育を抑制することは生存や資源の活用の観点から有利に働くものと考えられます。一方、農地の管理された栽培条件下では、植物のストレス応答による過剰な生育抑制が問題になる可能性があります。ストレスに応答した生育の調節機構の研究は、ストレス下での耐性と農業生産の両立を目指す上でも重要です。
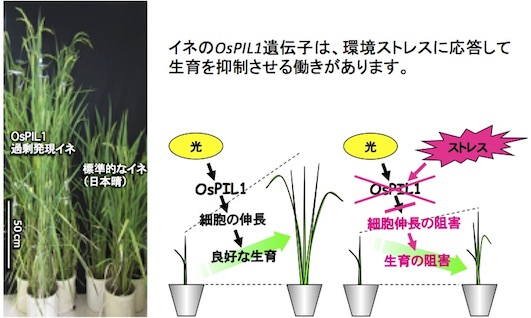
図7.OsPIL1は通常の生育条件でイネの伸長を促進しているが、ストレス環境下ではその発現が抑制され、結果的に細胞伸長が抑制される。このことがストレス下でのイネの生育阻害に関わっている (Todaka et al. 2012)。
(4)ストレス応答を利用した作物の形質制御
ストレスに応答して活性化される耐性機構は様々ですが、中には作物として有用な変化を伴う耐性機構もあります。例えば、トマトや果樹においては灌水制限や塩水を用いた浸透圧ストレスによって、糖度が高まることが知られています。また冬場に出回る寒締めホウレンソウは低温処理により糖度が高くなる特徴があります。これらは本来ストレス耐性を向上させるための植物の応答ですが、それが付加価値を向上させていることになります。また、ストレス応答で機能するDREB転写因子を導入すると、シロイヌナズナだけでなく、サトウキビでも糖の蓄積量が上がることが示されています。以上のような背景に基づき、ストレス応答やストレス応答関連遺伝子を利用して作物の形質制御を行う研究や、果実の高糖度化を題材としたストレス応答機構の研究を行っています。これらの研究の一部は産学連携の枠組みで行っています。
研究材料

モデル植物のシロイヌナズナ、また作物としてイネやトマトを用いてストレス応答の研究をしています。