地球温暖化の原因となる二酸化炭素等の温室効果ガスの排出を削減することは国際的に極めて重要な課題です。特に我が国は2005年発効した京都議定書により、2008年~2012年の二酸化炭素等の温室効果ガス排出量を1990年比で6%削減することが義務付けられており、そのための戦略の構築が求められています。その一つとして、持続可能な資源である植物バイオマスのエネルギー源としての利用は最も有望な手段であると考えられ、現在、米国、ブラジルなどを中心としてトウモロコシやサトウキビなどからバイオエタノールが生産され自動車燃料としての利用が進んでいます。しかし、食糧や飼料を利用することに対しては世界各地で問題となっており、今後、食糧と競合しないセルロース系バイオマスからのバイオエタノール生産に切り替える必要が指摘されています。
私たちは、麹菌を利用して、下記のようなセルロース系バイオマスの効率的な糖化に関する研究を進めています。
シロアリ共生系セルラーゼ遺伝子群の麹菌による大量発現系の構築とそれを用いた木質バイオマスの高度糖化・利用技術の開発
植物バイオマスの主要な成分である結晶性セルロースはその難分解性のため利用が進んでいませんが、自然界では全植物バイオマスの約20%はシロアリによって分解されていると言われ、特に腸内に原生生物が共生する「下等シロアリ」の場合、効率の良い種の場合はほぼ100%近いセルロースが資化され、リグニンのみが排出されることがわかっています。このことは、リグニン分解を介することなく非常に高効率でセルロースおよびヘミセルロースを資化する生化学的な系をシロアリが持っていることを意味しています。
理化学研究所および長崎大学のグループでは、シロアリが植物バイオマスを効率よく利用できるのはその腸内に多様な原生生物群を有しており、それらがセルロースを食胞内に取り込んだのち、独自の消化システムでグルコースにまで分解を行うためであることを明らかにしています(図1)。これまでに、シロアリ腸内に共生する原生生物由来のセルラーゼ・ヘミセルラーゼとして140以上の新規遺伝子が同定され、その機能解析が待たれています。しかし、これらの原生生物は培養ができないため、それら酵素群を利用可能な量を入手し、その性質を検討したり、実際にバイオマス分解に用いることは困難でした。
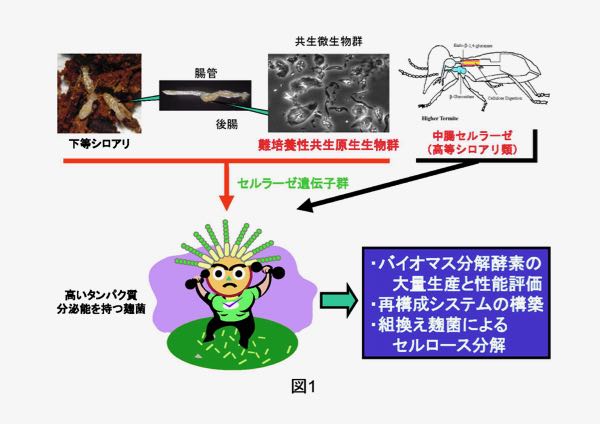
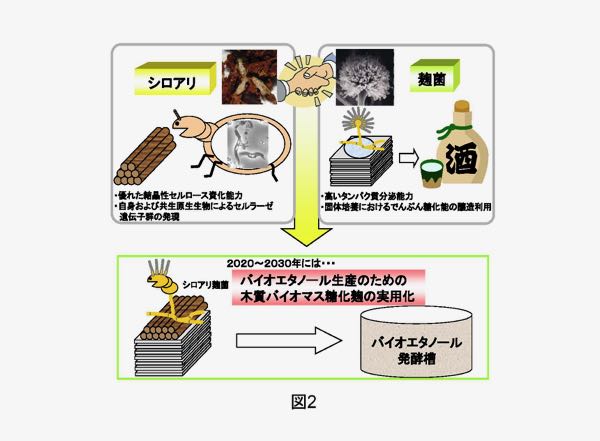
一方、農業生物資源研究所・琉球大学のグループは、シロアリ自身が生産するセルラーゼの研究を行っています。高等シロアリでは下等シロアリに見られるような腸内に共生する原生生物の存在は知られていないため、シロアリ自身のセルラーゼがセルロースの分解に中心的な役割を果たしていると考えられています(図1)。 当研究室では、上記4グループとの共同研究により、シロアリおよび腸内の原生生物群に由来するバイオマス分解酵素(セルラーゼ、ヘミセルラーゼなど)を、高いタンパク質分泌能を持つ麹菌で大量生産し、それを用いて植物バイオマスからグルコースやエタノールを生産する新しい技術を開発しようと考えています。将来的には、シロアリの持つバイオマス分解能力を再構成したり、それを麹菌に付与した「シロアリ麹菌(?)」(図2)の育種にもつなげてゆきたいと考えています。
セルロース緩和物質を高生産する麹菌の育種とバイオマス糖化への利用
主要な植物バイオマスとしてはデンプンとセルロースがあり、どちらもグルコースを重合した高分子化合物ですが、その結合形態に違いがあります。デンプンは植物の種子中に発芽のための栄養源として貯蔵されるものですので、必要なときにはアミラーゼにより容易にグルコースに分解される性質をもっています。一方、セルロースは植物体の骨格にあたる働きをもっており、分解されにくい堅固な構造をもっています。セルロースの分解にはセルラーゼを利用しますが、実際のプロセスにおいて大量のセルラーゼが必要であるという課題が指摘されています。一方で、植物が成長する過程においてセルロース繊維を緩める機能を有する“エクスパンシン”の存在が知られており、近年、これと相同性を有する因子として、糸状菌トリコデルマ・リーセイ由来の“スウォレニン”が単離されています。スウォレニンをセルラーゼに添加することにより、セルロース分解能を向上させることがすでに報告されています。私たちは、より効率的なセルロース分解システムの構築を目指して、これらのセルロース緩和物質を高生産する麹菌の育種も行っています。
将来的には、セルロース緩和物質と種々のセルラーゼを高生産する麹菌を育種し、固体培養による麹を製造し、平行復発酵によるバイオエタノールの生産を目指しています。
セルロース糖化麹菌の育種と固体培養によるバイオエタノール生産
麹菌は、日本酒の製造に長く使用されていることからもわかるように、米のデンプンを糖化する高い能力があります。デンプンを糖化するのに特化した微生物ともいえるでしょう。一方、セルロース分解能力は低いのですが、ゲノム解析の結果、多数のセルラーゼ遺伝子をもつことも明らかになりました。従って、麹菌も潜在的にはセルロース系バイオマスの分解能力をもっていることが期待されます。そこで、私たちは以下のようにして、セルロースを分解するのに特化した麹菌を育種し、米ではなく紙や木材などのセルロースを原料とした麹をつくり、清酒酵母で併行複発酵を行うことによりバイオエタノールを生産することを計画しています。 私たちは、セルロース系植物バイオマスの効率的な分解を目的として、高いセルラーゼ活性を示すAsprgillus aculeatusが生産する主要な3種類のセルラーゼ (FI-CMCase, cellobiohydrolase I, b-glucosidase 1)遺伝子を麹菌に導入することによって、上記のセルラーゼを同時に発現する3重発現株を作製しました。さらに、セルラーゼ3重発現株のセルロース資化能やセルロース分解能を向上させるため、セルラーゼやキシラナーゼ遺伝子の発現を制御する転写因子AoXlnRの高発現を試みています。これらのセルラーゼ3重発現株を用いて固体培養を行なうことによって、効率的なセルロース系植物バイオマスの分解システムの構築を試み、さらに、この固体培養物を用いて、セルロースからのバイオエタノールの生産を計画しています。