タンパク質の細胞内輸送は、主に輸送小胞を介して行われます。例えば分泌タンパク質や液胞内のタンパク質は、小胞体でフォールディング(折り畳み)や糖鎖付加などの修飾を受けた後、輸送小胞中に集められ、ゴルジ体へ運ばれます。ゴルジ体で更なる糖鎖付加やプロセッシングなどの修飾を受けたのち、分泌タンパク質と液胞タンパク質は別々の輸送小胞中に集められ、それぞれ細胞膜または液胞へと運ばれます。
麹菌は多種多様な酵素を菌体外に多量に分泌する性質をもつため、異種タンパク質生産の宿主として世界的に注目されています。このため麹菌の細胞内タンパク質輸送機構の解明は、産業的にも重要な意味を持っています。しかし麹菌を含め糸状菌全般において、タンパク質分泌が主に菌糸先端から起こるという知見をのぞくと、タンパク質の細胞内輸送に関してほとんどわかっていませんでした。
小胞体シャペロンを用いた麹菌の小胞体の可視化1, 2)
BiPは小胞体においてタンパク質のフォールディングを助けるシャペロンのひとつであり、麹菌ではbipA 遺伝子によりコードされています。当研究室では小胞体を可視化するため、BipA とEGFP(Enhanced Green Fluorescent Protein) との融合タンパク質を麹菌で発現させました。このBipA-EGFP 発現株を蛍光顕微鏡で観察したところ、核膜と思われる環状構造の他に、発達した網目状の蛍光像が観察されました(図1)。小胞体は英語でEndoplasmic Reticulum (ER)であり、直訳すると「細胞質中に広がる網目状構造」となりますが、まさにこれにふさわしい蛍光像といえます。また、経時的に観察することにより、この構造が活発に動いていることを明らかにしました。
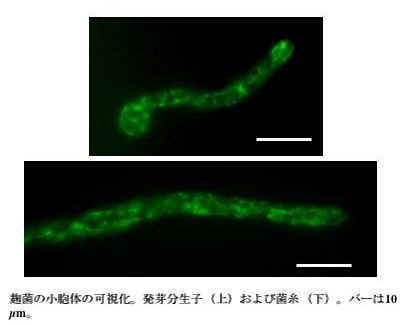
図1
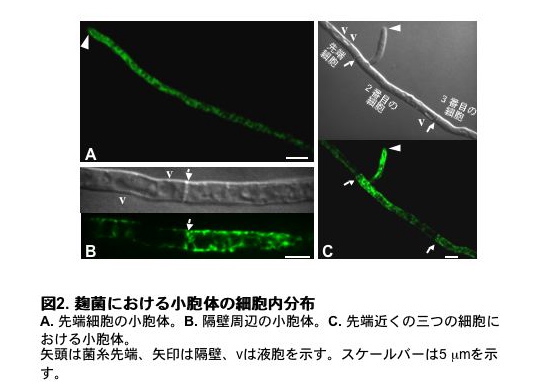
さらに詳しく観察したところ、個々の細胞において、小胞体は菌糸先端方向ほど密に存在し、菌糸を基部側にさかのぼるにつれてまばらになっていくことが分かりました(図2A, C)。このため細胞の仕切りにあたる隔壁を境にして、小胞体の分布は大きく異なります。隔壁の先端側では小胞体はあまり見られず、液胞が発達します。これに対して隔壁の基部側では、小胞体が発達します(図2B)。この結果は、隔壁によって隔てられた個々の細胞が、それぞれ独立して先端へ向かう極性を維持していることを意味しています。
麹菌のSNAREタンパク質の網羅的局在解析3, 4)
それでは他の小胞輸送に関わるオルガネラは、糸状菌細胞の中でどのような空間的配置をされているのでしょうか。私たちは輸送小胞が標的オルガネラと融合する際に働くSNAREと呼ばれる21個のタンパク質とEGFPの融合タンパク質を用いることで、小胞輸送に関わる様々なオルガネラを可視化しました。
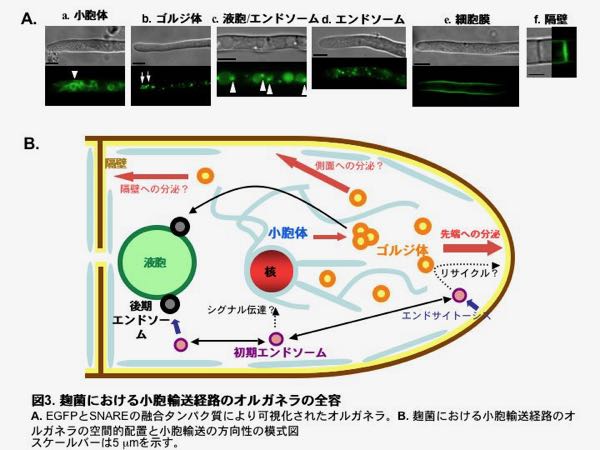
EGFPとSNAREの融合タンパク質により可視化された小胞体は、先に述べたBipAとEGFPの融合タンパク質で観察されたものとほぼ同様の分布を示しました(図3A-a)。また、ゴルジ体は菌糸先端近くの粒状構造体として観察されました(図3A-b)。分泌に関わる小胞体やゴルジ体が菌糸先端に密に存在していることは、菌糸先端からの分泌をサポートするために重要であると考えられます。
これに対して物質の分解に関わる液胞は菌糸先端にはあまり存在せず、菌糸を基部にさかのぼるにつれて数や体積が増す傾向にありました(図3A-c)。液胞近傍には粒状の構造があり、液胞へ輸送される物質の分別を行う後期エンドソームであることが分かりました(図3A-c)。また、菌糸の長軸方向に移動する粒状の構造体も見出されました。エンドサイトーシス経路の可視化の項で紹介している様に、これらは50 µmもの距離を平均5 µmの速度で動く初期エンドソームであることが分かりました(図3A-d)。
分泌の最終段階で働く細胞膜のSNAREタンパク質は細胞表層にほぼ一様に存在しましたが、意外なことに菌糸先端にはむしろ少なく存在しました (図3A-e)。また、細胞膜のSNAREは、細胞間の仕切りにあたる隔壁にも存在しました (図3A-f)。これまで、糸状菌において分泌は菌糸先端から起こるとされていましたが、細胞膜SNAREのこのような局在は、隔壁からも分泌が起こることを意味しているのかもしれません。
このように、小胞輸送のオルガネラは雑然と存在しているのではなく、細胞内の決まった場所に配置されていることが分かってきました(図3B)。麹菌は細胞内の領域をうまく使い分けることによって、先端生長などを効率よく行っていると考えられます。
RNase T1を用いた分泌タンパク質動態の解析5, 6)
私たちは麹菌の分泌酵素であるRNase T1とEGFP との融合タンパク質を発現することで、その挙動を観察しました。RNase T1-EGFP 融合タンパク質は、分泌酵素と同様に培養液中に分泌されます。蛍光顕微鏡による観察では、EGFP 蛍光は菌糸全体に分布しましたが、菌糸先端により明るく観察されました(図4)。この結果は、RNase T1-EGFP 融合タンパク質を運んでいる分泌小胞が伸長する菌糸細胞の先端に向かって流れていることを示唆しており、アミラーゼなどの酵素は菌糸先端から分泌されるとの従来からの説を支持するものです。一方、菌糸基部を観察したところ、RNase T1-EGFP 融合タンパク質は隔壁に多く局在しました(図5)。この結果は、先ほどのSNAREタンパク質の解析と同様に、タンパク質分泌が隔壁においても起こりうることを示唆しています。今後RNase T1-EGFP 融合タンパク質を用いることで、このような未知の分泌経路が実際に存在するかどうか調べていけるのではないかと考えています。
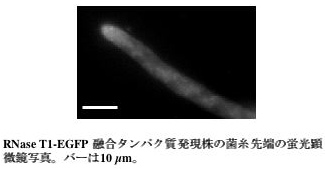
図4
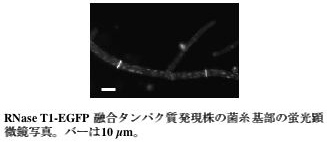
図5
参考文献
1) Maruyama et al., Fungal Genet. Biol., 2005
2) Maruyama et al., FEMS Microbiol. Lett., 2007
3) Kuratsu et al., Fungal Genet. Biol., 2007
4) Shoji et al., Biotechnol. Lette. 2008
5) Masai et al., Biosci. Biotechnol. Biochem., 2003
6) Masai et al., Biosci. Biotechnol. Biochem., 2004