液胞は植物や菌類の有する酸性コンパートメントであり、物質の貯蔵や分解、細胞質イオンの恒常性維持に重要なオルガネラです。糸状菌においては、液胞は古い細胞において発達していますが、生長先端近くの若い細胞においては未発達です。これは、液胞が長い糸状菌の菌糸の中で、場所によって様々な働きをしていることを意味しているのかもしれません。
CPYとEGFPの融合タンパク質を用いた麹菌液胞の可視化1)
私たちは液胞局在酵素カルボキシペプチダーゼY(CPY)とEGFP(Enhanced Green Fluorescent Protein)との融合タンパク質を麹菌において発現させることにより、麹菌の液胞を可視化することに成功しました。その結果、液胞は分生子内にも多く存在し(図1)、分生子の発芽において何らかの役割を果たしている可能性が示唆されました。また、これまでよく知られていた微分干渉像で凹んだ構造として観察される液胞のほかに、糸状菌特有のチューブ状、あるいはリング状の液胞様構造が存在することを明らかにしました(図1)。
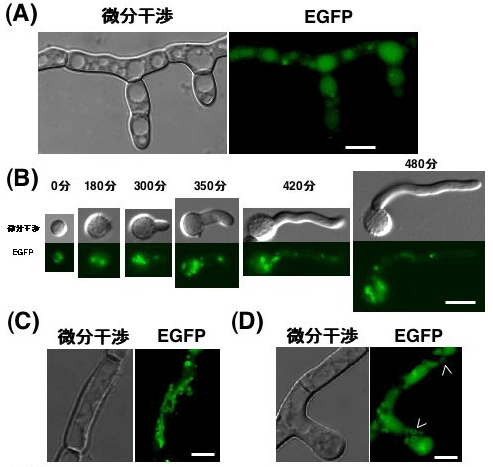
図1
麹菌におけるオートファジーの生理的意義の解析2, 3)
液胞はオートファジーと呼ばれる機構により細胞質成分を取り込み分解することで、飢餓時の栄養源リサイクルや細胞質の再構築にも働いています。私たちは上記のEGFPを付加したCPYを液胞のマーカーとして、DsRedを細胞質のマーカーとして用いることで、様々な培養条件下においてオートファジーによる細胞質の取り込みを検討しました。この結果、分生子の膨潤(図2A)、発芽(図2A, B)、分生子形成(図2C)など麹菌の各発達段階において、オートファジーによる細胞質成分の液胞への取り込みが起こっていることを見出しました。このように、液胞は糸状菌の分化においても重要な役割を果たしていることが分かりました。
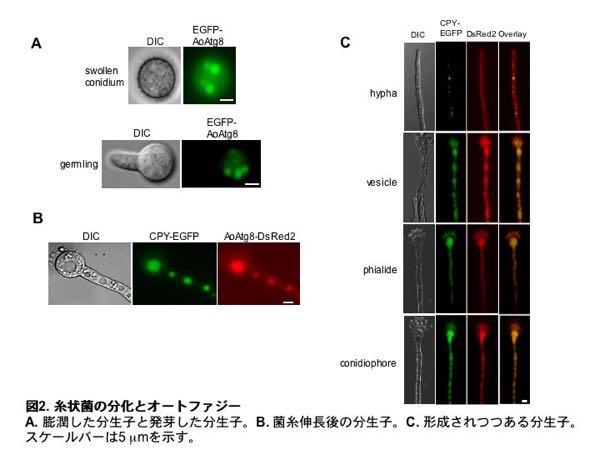
糸状菌の多様な液胞形態とその生理的意義の解析4)
また、麹菌の液胞膜上に存在するt-SNAREであるAoVam3pとEGFPとの融合タンパク質を用いることで液胞膜を可視化し、麹菌の液胞を詳細に観察することにも成功しました。共焦点レーザー顕微鏡により観察することで、液胞同士の融合、丸い液胞からのチューブ状液胞の形成、チューブ状液胞の動きなど、麹菌の液胞膜の動態を観察することができました(図3)。(動画1・動画2・動画3。動画は実際の25倍の速度で再生されています。)このように、糸状菌の液胞形態は極めて多様で、経時的に変化していくことが分かりました。
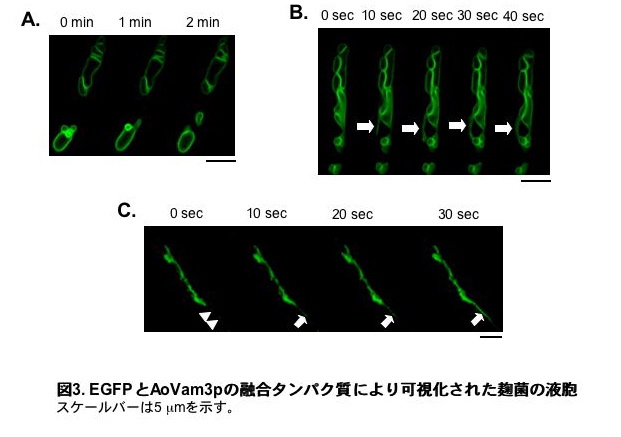
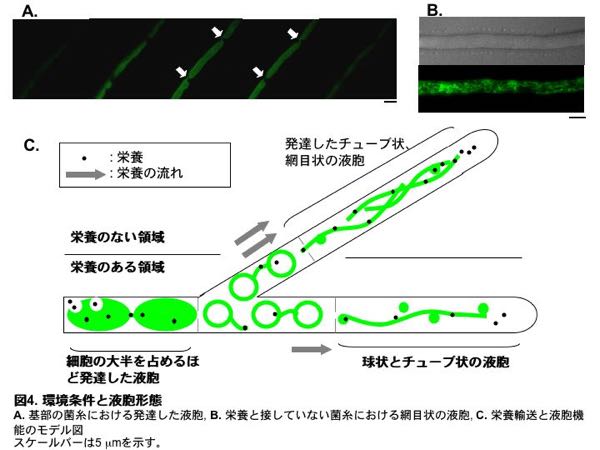
それでは液胞の多様な形態は、糸状菌の生育にどのような意義を持つのでしょうか。EGFPとAoVam3pの融合タンパク質を用いて基部(根元)の古い細胞の液胞を調べたところ、液胞が細胞体積の大半を占めるほど発達していることが分かりました(図4A)。さらに、オートファジーによる細胞質成分の取り込みと分解が、液胞体積増加の原因であると推測されました。また、これらの発達した液胞は栄養輸送に関わるとされるチューブ状液胞により連結されていました。このことから、細胞質成分の分解により生じたアミノ酸などは、チューブ状液胞を介して他の細胞に送られ栄養として再利用されていると考えられます。一方で、栄養と接していない細胞においてはチューブ状液胞が極めて発達し、網目状構造をとることも分かりました(図4B)。これらを総合して、増殖に寄与しない基部細胞の成分を液胞が分解し、それにより生じたアミノ酸などの栄養をチューブ状液胞により菌糸先端、特に栄養の足りない菌糸へと運んでいるのではないかという仮説を私たちは提示しています(図4C)5)。
麹菌の液胞形成に関するAovps24の機能解析6, 7)
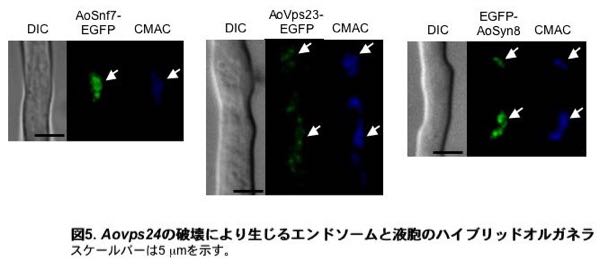
このように糸状菌の生育に重要な働きをしている液胞は、どのように形成されているのでしょうか。単細胞真核微生物の出芽酵母において、液胞を形成するタンパク質や膜は、小胞体、ゴルジ体を経た後エンドソームにおいて分別され、液胞に運ばれると考えられています。このとき、エンドソームでの分別に関わるタンパク質の一つとしてVps24というタンパク質が知られています。出芽酵母では、VPS24遺伝子の破壊によって液胞へのタンパク質輸送が異常になりますが、液胞形態や生育は大きな影響を受けません。ところが麹菌のVPS24相同遺伝子Aovps24を破壊したところ、液胞が細分化されるとともに麹菌の生育が著しく阻害されました。さらに細分化した液胞にはエンドソームのタンパク質が蓄積していたことから、Aovps24の破壊により液胞とエンドソーム両方の特徴をもつオルガネラが出現することが分かりました(図5)。このことから、麹菌では出芽酵母とは若干異なる機構で液胞が形成されていることが分かりました。
麹菌のエンドサイトーシス経路の可視化8)
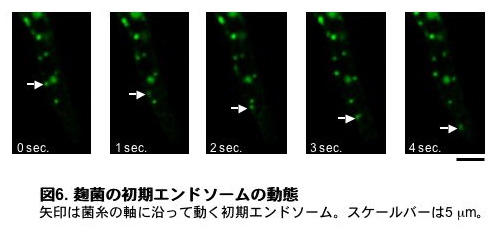
液胞はエンドサイトーシスと呼ばれる機構により取り込まれた細胞外や細胞膜上の物質を分解することで、細胞の環境適応や栄養分の取得に働いています。しかし、糸状菌においてはエンドサイトーシスの存在自体が長い間疑問視されていました。私たちは細胞膜上の推定プリントランスポーターであるAoUapCとEGFPの融合タンパク質を用いることで、麹菌においてエンドサイトーシスが起こるかどうかを調べました。その結果、培地へアンモニウム塩を添加することによって、AoUapCとEGFPの融合タンパク質がエンドサイトーシスにより細胞内へ取込まれることを見出しました。さらにエンドサイトーシスによる取り込みに際して、細胞内に粒状のエンドソーム構造が観察されるようになりました。これらエンドソーム構造は平均約5 µm/secの速さで、菌糸内を長軸方向に微小管依存的に移動し、その移動距離が50 µmを越えるものも観察されました(図6、 動画4、動画は実際の速度で再生されています)。このように、糸状菌でも酵母や植物、動物と同様にエンドサイトーシスが起こっていることが明らかになりました。しかし、なぜエンドソームがこれほどの長距離を高速で移動する必要があるのか、その生理的意義の解明は今後の課題として残されています。
これまで真核微生物のモデルである出芽酵母においては、液胞研究が盛んに行われてきました。しかし上記のように、糸状菌にはチューブ状液胞や細胞ごとに形態の違う液胞といった、出芽酵母には見られない特徴が存在することが分かってきました。このような特徴は、顕著な極性を持つ多細胞生物として糸状菌が生きるため、進化の過程で獲得されたものと考えられます。
植物との共生、タンパク質の高分泌といった地球環境や人間生活に重要な糸状菌の働きの多くは、出芽酵母には見られないものです。液胞研究を通して見えてきた糸状菌の独自性は、このような出芽酵母には見られない糸状菌の働きをサポートしているのではないでしょうか。つまり、糸状菌独自の生理現象を明らかにしていくことが、糸状菌の生態系における働きの理解と、それを私たちの生活に役立つよう応用することにつながるのではないかと私たちは考えています。
参考文献
1) Ohneda et al., Fungal Genet. Biol., 2002
2) Kikuma et al., Eukaryot. Cell, 2006
3) Kikuma et al., Autophagy, 2007
4) Shoji et al., Eukaryot. Cell, 2006
5) Shoji et al., Autophagy, 2006
6) Tatsumi et al., Biochem. Biophys. Res. Commun., 2006
7) Tatsumi et al., Biochem. Biophys. Res. Commun., 2007
8) Higuchi et al., Biochem. Biophys. Res. Commun., 2006