植物の栄養応答経路の解析
植物の栄養環境に対する応答反応のしくみを調べる。
植物が無機養分だけで生育することができるといっても、ほとんどの土壌では、養分はわずかしか含まれていません。 私たちが空腹を感じて食事をするのと同様に、植物も栄養分が少ないことを感知して、その栄養分を効率良く吸収したり、利用したりする仕組みを備えています。
例えば、硫酸のトランスポーターは硫黄が欠乏するとmRNAレベルで発現誘導を受けます。この過程に重要な代謝産物を同定したり(Kim, H. et al., 1999, Planta)、その制御機構を明らかにしています(Ohkama-Ohtsu, N. et al., 2004, Plant Phys)。
また、ホウ素トランスポーターはホウ素栄養条件に応じてそのタンパク質の蓄積量がエンドサイトーシスを介して制御されることを明らかにしています (Takano et. al., 2005, PNAS)。最近では、ホウ素トランスポーターが種類によって根の細胞の土壌側に面した細胞膜と、導管に面した細胞膜に局在して、細胞を横切るホウ素の輸送を担っていることを明らかにしました(Takano et al., 2010, PNAS)。このような局在や分解に関与するアミノ酸を同定したりしています(Kasai et al 2011 JBC)シロイヌナズナとイネでのホウ素トランスポーターの役割や制御後外も明らかにしています(Nakagawa et al 2007 Plant Cell)。
最近では、ホウ素トランスポーターのホウ素条件に応じた発現制御に5’UTR配列を介したmRNA分解が関与していることを明らかにしており(Tanaka et al 2011 Plant Cell)、リボソームを介した栄養感知機構の可能性を追求しています。
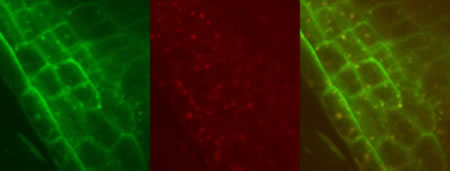
Confocal image of a root tip of transgenic Arabidopsis plants demonstrating the pathway of plasma membrane protein degradation via endosome. A boron transporter BOR1 fused with GFP (green) was internalized from plasma membrane and localized transiently in endosomes labelled with a Rab-GTPase Ara7 fused with mRFP (red) and subsequently degraded in vacuoles upon high-boron supply.