細胞運動について
細胞運動のテーマでは、微生物を含む動物型の真核細胞について、細胞質分裂、細胞移動(遊走)、エンドサイトーシス(特に貪食作用とマクロピノサイトーシス)といった広い意味での細胞運動現象とその基盤となる細胞基質間接着の分子メカニズムを解明しています。これらの現象で、細胞はその形態を厳密な時間的・空間的制御のもと変化させますが、その制御された動きを生み出す分子メカニズムを細胞性粘菌のアメーバ細胞を用いて調べています。細胞壁を持たない細胞の形態はアクチンや微小管といった細胞骨格系により決まるので、細胞運動の研究は細胞骨格の再構築(reorganization)機構の研究ともいえます。実験材料に用いている細胞性粘菌については下記(モデル生物:細胞性粘菌)をご参照ください。以下、細胞性粘菌を単に粘菌と呼びます。
細胞運動に関する研究テーマ
1)動物型細胞質分裂の分子メカニズム
粘菌のアメーバ細胞は、動物培養細胞と形態的に大変良く似た細胞質分裂を行います。1987年には、細胞性粘菌のミオシンII(筋肉のミオシンと同じタイプのミオシン)の遺伝子破壊によって細胞が多核化することが示され、このことによって、動物型真核細胞の細胞質分裂にミオシンIIが関与することの証明に粘菌が貢献するとともに、粘菌が高等動物まで普遍的な細胞質分裂の分子メカニズムを持つこともわかりました。当グループは、世界で初めてタギング法(REMI法)により多核細胞を生じる粘菌変異株を多数分離し、そのうちの1株の解析からIQGAP様タンパク質が細胞質分裂の終結に関わることを他の生物に先駆けて証明しました。現在では、相互作用するタンパク質の探索などIQGAP様タンパク質に関する研究に加え、別の変異株から同定したMKLP1様タンパク質(cytokinesin/kif12)、新規タンパク質CykA、CykB、D47-1p、D411-2pの細胞質分裂における役割を解明するための研究を進めています。粘菌を用いて細胞質分裂に普遍的に関わる新規分子を同定し、その機能を他の生物に先駆けて明らかにすることを目指しています。当グループが最も力を入れているテーマです。
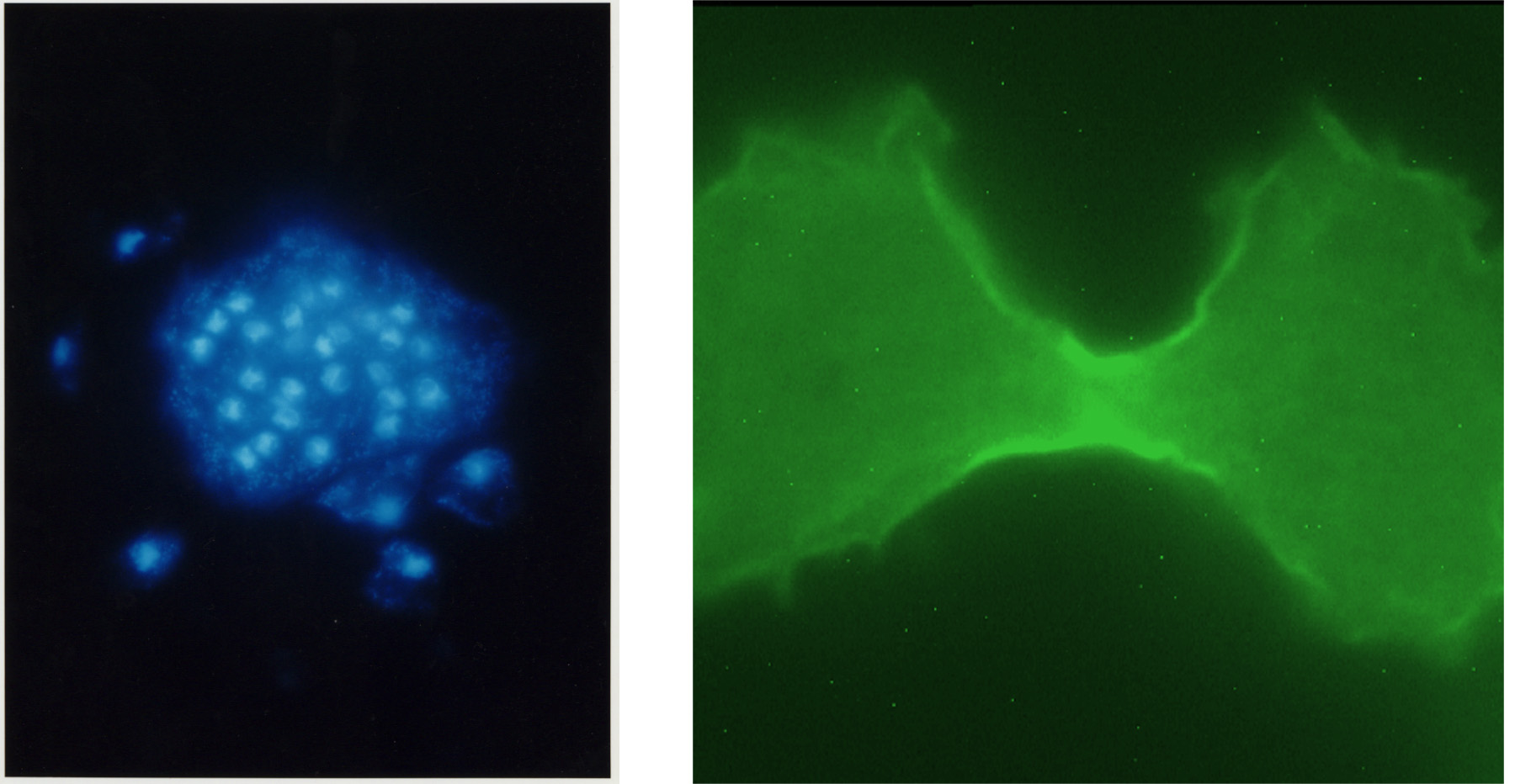
(図の説明)左は、粘菌の細胞質分裂変異株(IQGAP様タンパク質の遺伝子破壊株)の巨大多核細胞の核をDAPI染色したもの。右はGFPを融合したIQGAP様タンパク質の蛍光像で、IQGAP様タンパク質が細胞の表層を細胞質分裂時にできるくびれ(分裂溝)に向かって集まってきたところ。
2)RhoファミリーGTPaseの機能解析
RhoファミリーGTPaseは低分子量GTPaseのサブファミリーで、細胞骨格の再構築を通じて細胞質分裂、貪食作用、細胞移動(遊走)などの細胞運動現象を制御する情報伝達系に関わることが知られています。また、Rhoファミリーには、その活性を調節することで情報伝達に関わるRhoGAP、RhoGEF、RhoGDIがあります。細胞性粘菌には少なくとも15種類のRhoファミリーが存在し、RacEのように関与する機能(細胞質分裂)がわかっているものもありますが、多くは機能未知です。また、GAPやGEFはさらに種類も多く、そのほとんどが機能未知です。当グループは、これらRhoファミリーとその制御分子群が等グループが注目している細胞運動現象に関わるか,また、どのように関わるかを標的遺伝子破壊と分子の細胞内局在を手掛かりに明らかにしようとしています。これまで、RhoGDIのひとつであるRhoGDI1が細胞質分裂に関わること、RhoGDI1がどのRhoファミリーを制御するかを明らかにしましたが、その他の分子についてもRhoファミリー自身を中心に解析中です。
3)細胞基質間接着の分子メカニズム
細胞の遊走に基質接着は必須で、その動的変化により細胞は基質に対して動きます。さらに基質接着は細胞質分裂とも切り離せない関係で、粘菌では接着に依存した細胞質分裂機構もあるようです。動物では細胞は細胞外マトリックスと接着斑を通じて接着しますが、接着斑を構成する分子の粘菌オルソログや基質接着変異株を利用して基質接着のメカニズムだけでなく、細胞質分裂、細胞遊走との関わりも明らかにするプロジェクトで、最近力を入れています。
4)細胞遊走、貪食作用、マクロピノサイトーシスの分子メカニズム
D411-2pのように、細胞質分裂に関わるタンパク質を同定すると、しばしばそれが細胞遊走、貪食作用、マクロピノサイトーシスといった細胞骨格の再構築を含む現象にも関わることがあります。そのような分子では、細胞質分裂に加えてこれらの現象における役割を明らかにする実験も行っています。そのような解析から、細胞骨格の再構築における現象をこえた機能を明らかにすることができるかもしれません。
モデル生物:細胞性粘菌
細胞運動の研究に用いている細胞性粘菌Dictyostelium discoideumは真核の微生物です。自然界では森の下生えなどに生息し、細菌を餌にして増殖する単細胞アメーバです。このアメーバ細胞は、細胞壁を持たず基質に接着して二分裂で増殖し、餌である細菌に走化性運動により近づき、貪食作用によりこれを取込むので、形態も含めて動物の白血球や培養細胞に大変良く似ています。最近の研究から、細胞性粘菌は形だけではなく細胞質分裂や貪食作用のメカニズムが動物と分子レベルまで似ていることがわかってきたので、こうした動物型細胞運動現象を解析するモデル生物として注目されています。また、餌を食い尽くして栄養がなくなると、10万個ほどのアメーバ細胞がcAMPに対する走化性運動により集合して多細胞体を形成し、それが飢餓開始から約1日の間に休眠胞子をもつ子実体へと劇的に変化するので、走化性運動のメカニズムの解析に加え、発生生物学では細胞分化と形態形成のモデル生物としても用いられています。
ゲノムサイズは34M塩基対と酵母の2倍強ほどで、無性世代では半数体のまま上記の様々な生命現象を示します。相同組換えによる標的遺伝子破壊とタギング法(REMI法)が確立しているので、変異株とその表現型から遺伝子と生理機能の関係を明らかにすることができます。これは、同じ真核細胞の酵母(出芽酵母と分裂酵母)が得意とするところですが、酵母は細胞壁を持っているため素早く形を変えられず、基質にも接着できないため、上記動物型細胞運動現象そのものが存在しません。酵母で研究できることは酵母に任せた方が良いのは明らかですが、細胞の形を素早く変化させて引き起こす細胞運動現象は、細胞性粘菌で研究するのが一番よいのです。ゲノムの全塩基配列は2005年に報告され、データベースDictyBaseからあらゆる情報や実験材料を引き出すことができるようになっています。細胞性粘菌に関する学会も、国際集会(DictyBase参照)、日本の細胞性粘菌研究会とも毎年開かれており、活発な議論がなされています。